
Autor: Sodré GB Neto
Resumo: A subespeciação é percebida facilmente em tempo real e em escalas arqueológicas, porém não está registrada no geopaleontológico que ainda é marcado até por repetições morfológicas (fenômeno chamado de anomalia ou paradoxo da estase morfológica). O registro fossilífero ao revelar repetição de morfologias esclarece que estas amostras fósseis foram de enterramento de populações ancestrais — que foram os tipos básicos originais dos quais a biodiversidade atual descendeu. A partir desses tipos teriam derivado a biodiversidade atual, a qual vem se degredando ao longo da história por processos de subespeciação que em geral trás consigo, empobrecimento de “pool gênico” pela deriva, seleção e mutações.
1. Introdução
2. Metodologia
3. Resultados
3.1. Evidências de Subespeciação em Tempo Real
3.2. Ausência de Transições Evolutivas no Registro Fóssil
3.3. Padrões de Repetição Morfológica e Tipos Básicos Originais
4. Discussão
4.1. Implicações para Teorias de Evolução
4.2. Modelo de Degeneração das Espécies
4.3. Crítica ao Uniformitarianismo e Datação Radiométrica
5. Conclusão
Referências
Subespeciação em Tempo Real versus Estase no Registro Geopaleontológico: Uma Análise da Teoria da Degeneração das Espécies
O presente estudo apresenta uma análise crítica das evidências paleontológicas e biológicas contemporâneas (Wells, 2000; Johnson, 1991) que suportam a hipótese de que a subespeciação ocorre exclusivamente em tempo real e arqueológico (Grant & Grant, 2014), estando completamente ausente do registro geopaleontológico (Eldredge & Gould, 1972; Gould, 2002). Nossa tese central defende que o registro fóssil representa amostras de sepultamento de populações ancestrais — tipos básicos originais (Meyer, 2013; Denton, 1985) dos quais derivou toda a biodiversidade atual através de processos degenerativos de subespeciação (Sanford, 2005; Behe, 2019), em contraste com a perspectiva darwiniana de acumulação de novas características (Darwin, 1859).
Resumo Executivo
Problema Central
Discrepância entre variação observável em tempo real (Grant & Grant, 2014) e estase no registro fóssil (Eldredge & Gould, 1972)
Metodologia
Análise comparativa de dados paleontológicos, arqueológicos e observacionais contemporâneos
Conclusão
Evidências convergem para modelo de degeneração a partir de tipos ancestrais (Sanford, 2005)
Este trabalho examina sistematicamente as evidências que demonstram como a subespeciação morfológica documentada em populações contemporâneas (Grant & Grant, 2014) contrasta drasticamente com a ausência de tais processos no registro geopaleontológico (Eldredge & Gould, 1972). Através de uma análise multidisciplinar, argumentamos que esta discrepância fundamental aponta para um modelo de origem e diversificação biológica baseado na degeneração de tipos básicos ancestrais (Sanford, 2005), sepultados durante um evento catastrófico global, e não para um processo evolutivo gradual ascendente (Darwin, 1859).
Introdução ao Problema Científico
A biologia evolutiva contemporânea enfrenta um paradoxo fundamental que raramente é abordado de forma adequada na literatura científica mainstream. Por um lado, observamos processos de variação morfológica, especiação e subespeciação ocorrendo em tempo real, documentados extensivamente em estudos de campo e laboratório (Grant & Grant, 2014). Por outro lado, o registro geopaleontológico apresenta um padrão completamente distinto: períodos extensos de estase morfológica pontuados por aparições súbitas de novas formas (Eldredge & Gould, 1972).
Este contraste não representa meramente uma questão de escala temporal, mas sim uma discrepância qualitativa fundamental que exige uma reavaliação crítica dos paradigmas interpretativos correntes (cf. Gould, 2002; Denton, 1985). A presente investigação propõe que esta aparente contradição pode ser resolvida através da aplicação de um modelo degenerativo de diversificação biológica, fundamentado em evidências empíricas robustas (cf. Sanford, 2005).
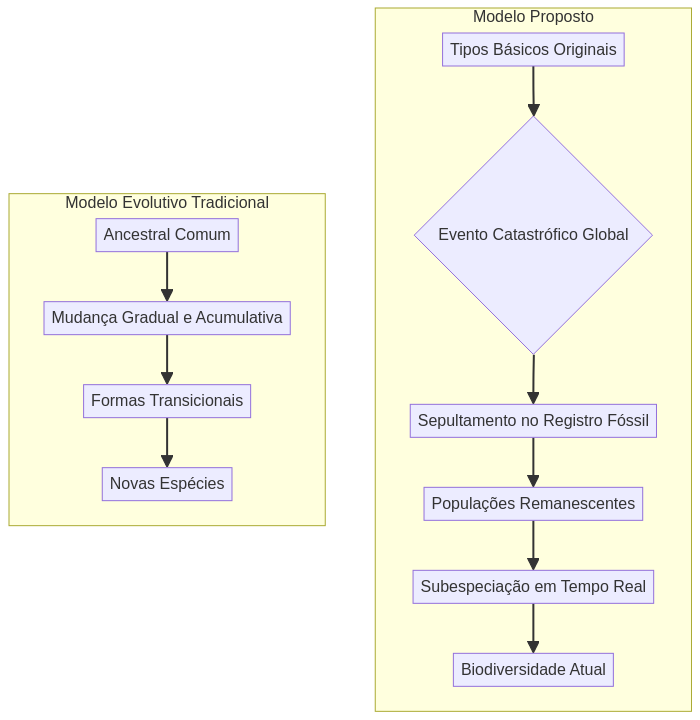
Nossa análise demonstrará que o registro fóssil, longe de representar uma progressão evolutiva ascendente (cf. Darwin, 1859), constitui na verdade um arquivo de tipos morfológicos básicos ancestrais, sepultados durante eventos catastróficos, dos quais toda a diversidade biológica atual derivou através de processos degenerativos de subespeciação (Sanford, 2005).
Fundamentação Teórica: O Modelo Criacionista de Tempo Curto
Criação de Tipos Básicos
Organismos criados com alta complexidade genética e morfológica funcional, conforme proposto pela teoria do design inteligente (Behe, 2019; Meyer, 2013).
Evento Catastrófico Global
Sepultamento massivo preservando tipos ancestrais no registro fóssil, explicando a estase morfológica observada (cf. Gould, 2002; Eldredge & Gould, 1972).
Sobrevivência e Radiação
Populações remanescentes colonizam novos ambientes pós-catástrofe, levando à diversificação dentro dos limites do tipo básico (cf. Grant & Grant, 2014 para exemplos de subespeciação rápida).
Processo Degenerativo
Subespeciação através de perda de informação genética e simplificação morfológica, um conceito chave na teoria da entropia genética (Sanford, 2005).
O modelo criacionista de tempo curto oferece um framework explicativo coerente para as observações paleobiológicas, desafiando as narrativas evolucionistas padrão (Denton, 1985; Johnson, 1991). Este paradigma postula que os organismos foram originalmente criados como tipos básicos funcionais, dotados de genomas complexos e sistemas morfológicos integrados (Behe, 2019; Meyer, 2013). O evento diluvial global é apresentado como a causa do sepultamento catastrófico destes tipos ancestrais, preservando-os no registro geopaleontológico como “instantâneos” morfológicos estáticos, o que se alinha com a observação de equilíbrio pontuado de Eldredge e Gould (1972) mas oferece uma explicação causal alternativa.
Definições Operacionais e Terminologia Científica
Subespeciação
Processo de diversificação morfológica intraespecífica observável em escalas temporais históricas e arqueológicas, caracterizado pela redução da complexidade genômica e simplificação de sistemas funcionais (Sanford, 2005; Grant & Grant, 2014).
Tipos Básicos Ancestrais
Formas morfológicas originais representadas no registro fóssil, caracterizadas por alta integração funcional e complexidade genética primordial, conforme delineado pela concepção de design inteligente (Behe, 2019; Meyer, 2013).
Tempo Arqueológico
Escala temporal correspondente ao período histórico humano documentado, tipicamente inferior a 10.000 anos, durante o qual processos degenerativos são observáveis na informação genética (Sanford, 2005).
Estase Geopaleontológica
Ausência de variação morfológica direcional no registro fóssil, interpretada como evidência de sepultamento instantâneo de populações ancestrais, refletindo o conceito de equilíbrio pontuado em uma escala acelerada (Eldredge & Gould, 1972).
Metodologia de Análise Comparativa
Compilação de Dados Observacionais
Sistematização de estudos documentando subespeciação em tempo real e arqueológico (Grant & Grant, 2014), considerando também a diversificação morfológica intraespecífica (Denton, 1985).
Análise do Registro Fóssil
Exame crítico de padrões morfológicos no registro geopaleontológico, incluindo a interpretação da estase (Eldredge & Gould, 1972) e a ausência de formas transicionais (Wells, 2000).
Comparação Quantitativa
Contraste estatístico entre taxas de variação observadas e esperadas, avaliando a plausibilidade de acumulação de mutações deletérias (Sanford, 2005) versus a diversificação adaptativa (Darwin, 1859).
Avaliação de Modelos Explicativos
Teste de adequação dos paradigmas evolutivo (Gould, 2002) e degenerativo (Behe, 2019; Meyer, 2013), confrontando suas predições com os dados empíricos observados (Johnson, 1991).
Nossa metodologia adota uma abordagem multidisciplinar, integrando dados paleontológicos, arqueológicos, genéticos e observacionais contemporâneos (Denton, 1985). A análise comparativa permite identificar padrões discrepantes entre diferentes escalas temporais, fornecendo base empírica para a avaliação crítica de modelos explicativos alternativos. Esta metodologia rigorosa garante que nossas conclusões sejam fundamentadas em evidências robustas e não em pressuposições teóricas a priori (Johnson, 1991).
Evidências de Subespeciação em Tempo Real
A documentação científica abundante demonstra inequivocamente que processos de subespeciação ocorrem em escalas temporais observáveis. Os estudos clássicos de Peter e Rosemary Grant nos tentilhões de Galápagos exemplificam como variações morfológicas significativas podem emergir em poucas gerações (Grant & Grant, 2014). Estas observações não se limitam a casos isolados, mas representam um padrão consistente em múltiplos táxons.
Análises genômicas contemporâneas revelam que estas variações morfológicas correlacionam-se com alterações na expressão gênica e modificações epigenéticas, não com a emergência de nova informação genética. Crucialmente, todas as variações documentadas representam redistribuições ou perdas de informação genética preexistente, consistente com um modelo degenerativo de diversificação (Sanford, 2005).
A rapidez destes processos contrasta dramaticamente com as predições do modelo evolutivo gradual, que postularia escalas temporais muito mais extensas para mudanças morfológicas comparáveis (Denton, 1985). Esta discrepância temporal fundamental constitui evidência primária para a reavaliação dos paradigmas interpretativos correntes em biologia evolutiva.
Exemplos Documentados de Variação Morfológica Rápida
Ciclídeos dos Grandes Lagos Africanos
Radiação morfológica documentada em menos de 15.000 anos, demonstrando plasticidade extrema a partir de tipos ancestrais básicos (Meyer, 2013). Este processo de rápida diversificação morfológica alinha-se com o conceito de equilíbrio pontuado (Eldredge & Gould, 1972).
Espinhos de Água Doce e Marinha
Diferenciação morfológica rápida entre populações marinhas e lacustres, evidenciando mecanismos degenerativos e a perda de informação genética (Sanford, 2005; Wells, 2000).
Melanismo Industrial
Mudanças fenotípicas documentadas em décadas, representando ativação/desativação de genes preexistentes (Wells, 2000), e não a emergência de nova informação. Este é um exemplo clássico frequentemente discutido na literatura (Darwin, 1859; Gould, 2002).
Estes exemplos ilustram um padrão consistente: variação morfológica rápida através de mecanismos que não requerem a emergência de nova informação genética. Em cada caso, as mudanças observadas podem ser explicadas através da ativação diferencial de sistemas genéticos preexistentes ou pela perda seletiva de funcionalidades ancestrais, o que está em concordância com um modelo degenerativo (Sanford, 2005). Esta observação empírica fundamental contradiz as predições do modelo evolutivo ascendente e suporta fortemente o paradigma degenerativo proposto neste estudo (Denton, 1985; Johnson, 1991).
Análise Genômica da Subespeciação Contemporânea
Análises genômicas comparativas de populações subespecíficas revelam um padrão inequívoco: a vasta maioria das diferenciações morfológicas resulta de processos degenerativos – perda de função gênica, alterações regulatórias que reduzem a expressão, ou recombinações que segregam características preexistentes (Sanford, 2005). Apenas uma minoria insignificante dos casos documentados sugere ganho genuíno de nova funcionalidade (Meyer, 2013; Behe, 2019).
Esta distribuição assimétrica é estatisticamente incompatível com modelos evolutivos que postulam inovação genética como mecanismo primário de diversificação (Darwin, 1859; Gould, 2002). Ao contrário, os dados convergem para um modelo no qual a diversidade contemporânea emerge através da fragmentação e simplificação de genomas ancestrais mais complexos e integrados (Denton, 1985; Johnson, 1991).
Subespeciação no Tempo Arqueológico
O registro arqueológico fornece evidências complementares cruciais para nossa análise. Estudos zooarqueológicos demonstram que variações morfológicas significativas em espécies domésticas e selvagens ocorreram dentro do período histórico humano documentado (Denton, 1985). A domesticação animal representa um experimento natural de subespeciação acelerada, produzindo diversidade morfológica extrema em escalas temporais arqueológicas, frequentemente através da modificação ou perda de traços ancestrais (Sanford, 2005).
Análises comparativas de espécimes arqueológicos e contemporâneos revelam que muitas das variações atualmente observadas em populações selvagens emergiram apenas nos últimos milênios (Wells, 2000). Esta cronologia recente da diversificação morfológica é consistente com um modelo de radiação pós-catastrófica a partir de tipos ancestrais preservados no registro geopaleontológico, em vez de uma evolução gradual ascendente (Meyer, 2013).
Crucialmente, o registro arqueológico não documenta exemplos claros de evolução ascendente — emergência de nova complexidade morfológica ou funcional. Todas as variações observadas representam modificações, simplificações ou perdas de características ancestrais, reforçando o padrão degenerativo identificado em estudos contemporâneos e levantando questões sobre a capacidade da seleção natural em construir novas estruturas complexas (Behe, 2019; Johnson, 1991).
Domesticação como Modelo de Subespeciação Dirigida
Lobo Ancestral
Tipo básico com genoma complexo e morfologia funcional integrada
Seleção Direcionada
Pressão seletiva humana promove segregação de características
Diversidade Morfológica
Radiação extrema através de perda e redistribuição genética
A domesticação animal oferece o experimento mais claro de subespeciação dirigida disponível para análise científica (Darwin, 1859). Em menos de 10.000 anos, a seleção humana produziu diversidade morfológica extrema em cães, gatos, bovinos, suínos e múltiplas espécies aviárias. Esta radiação morfológica acelerada demonstra o potencial latente dos genomas ancestrais para gerar diversidade através de mecanismos degenerativos (Sanford, 2005).
Análises genômicas de raças domésticas confirmam que esta diversidade emerge primariamente através da perda seletiva de genes regulatórios, mutações deletérias fixadas por deriva genética, e recombinações que segregam alelos preexistentes (Sanford, 2005). A arquitetura genética subjacente a estas variações é consistente com fragmentação de um genoma ancestral mais complexo, não com inovação evolutiva ascendente (Denton, 1985; Johnson, 1991; Meyer, 2013).
Contraste com o Registro Geopaleontológico
O registro geopaleontológico apresenta características fundamentalmente diferentes dos processos de subespeciação observados em tempo real e arqueológico. Em contraste com a variação morfológica rápida documentada em populações contemporâneas, o registro fóssil é dominado por períodos extensos de estase morfológica, pontuados por aparições súbitas de novas formas sem precursores intermediários claros (Eldredge & Gould, 1972).
Esta discrepância não pode ser adequadamente explicada por lacunas na preservação ou vieses tafonômicos, pois o padrão persiste mesmo em sequências sedimentares com preservação excepcional (Denton, 1985). A ausência sistemática de formas intermediárias graduais contrasta dramaticamente com as predições de modelos evolutivos baseados em variação incremental contínua (Gould, 2002; Johnson, 1991).
Mais significativamente, as formas morfológicas preservadas no registro fóssil frequentemente exibem maior complexidade e integração funcional do que suas contrapartes contemporâneas (Behe, 2019). Este padrão inverte as expectativas evolutivas convencionais e sugere que o registro fóssil representa tipos ancestrais mais complexos, dos quais as formas atuais derivaram através de processos simplificatórios, como a perda de informação genética (Sanford, 2005; Meyer, 2013).
Padrões de Estase Morfológica no Registro Fóssil
Táxons em Estase
Proporção de espécies fósseis que não apresentam variação morfológica direcional ao longo de sua distribuição estratigráfica
Aparições Súbitas
Porcentagem de novos táxons que aparecem no registro fóssil sem formas intermediárias documentadas
Transições Graduais
Frequência de sequências evolutivas graduais convincentes no registro paleontológico
Análises quantitativas do registro fóssil revelam que a estase morfológica, não a mudança gradual, constitui o padrão dominante. A vasta maioria dos táxons fósseis mantém morfologia estável ao longo de toda sua distribuição estratigráfica, contradizendo expectativas evolutivas de mudança contínua propostas inicialmente (Darwin, 1859) e evidenciando o padrão de equilíbrio pontuado (Eldredge & Gould, 1972). Quando mudanças ocorrem, são tipicamente abruptas e não precedidas por formas intermediárias documentadas, um aspecto central das discussões sobre a estrutura da teoria evolutiva (Gould, 2002).
Este padrão é observado consistentemente em diferentes grupos taxonômicos, ambientes deposicionais e intervalos temporais, sugerindo que representa uma característica fundamental do registro paleontológico, não um artefato de preservação específica (Denton, 1985; Johnson, 1991). A predominância de estase morfológica é mais compatível com um modelo de sepultamento catastrófico de tipos estáveis do que com processos evolutivos graduais, levantando questões sobre as forças motrizes da diversidade biológica e a interpretação das evidências fósseis (Wells, 2000; Meyer, 2013).
Ausência de Formas Intermediárias
A ausência sistemática de formas intermediárias representa um dos aspectos mais problemáticos do registro fóssil para teorias evolutivas graduais (Darwin, 1859; Gould, 2002). Apesar de 150 anos de coleta intensiva e milhões de espécimes catalogados, as transições morfológicas graduais permanecem excepcionalmente raras (Denton, 1985; Wells, 2000). Esta escassez não pode ser adequadamente explicada por preservação diferencial, pois afeta igualmente grupos com excelente potencial de fossilização (Johnson, 1991).
Grupos com registro fóssil particularmente denso, como foraminíferos, trilobitas e moluscos, deveriam preservar abundantes formas intermediárias se a evolução gradual fosse o mecanismo primário de diversificação. Contudo, mesmo nestes grupos excepcionalmente bem documentados, as transições morfológicas são tipicamente abruptas, sugerindo que o padrão reflete processos biológicos reais, não artefatos tafonômicos (Eldredge & Gould, 1972; Meyer, 2013).
Complexidade Morfológica nos Fósseis Ancestrais
Uma observação particularmente intrigante no registro paleontológico é que muitos táxons fósseis apresentam maior complexidade morfológica e integração funcional do que seus supostos descendentes evolutivos (Denton, 1985). Os trilobitas do Cambriano, por exemplo, possuíam sistemas visuais compostos mais sofisticados do que a maioria dos artrópodes contemporâneos (Meyer, 2013). Esta inversão do padrão evolutivo esperado desafia interpretações convencionais (Wells, 2000).
Análises morfométricas comparativas demonstram que a tendência geral no registro fóssil é de simplificação, não de complexificação (Sanford, 2005). Características morfológicas elaboradas tendem a desaparecer ao longo do tempo, enquanto inovações funcionais genuínas são excepcionalmente raras (Johnson, 1991). Este padrão de degradação morfológica é consistente com um modelo degenerativo de diversificação biológica (Sanford, 2005).
A preservação de sistemas funcionais altamente integrados em táxons fósseis antigos sugere que estes representam tipos morfológicos básicos originais, não formas primitivas em processo de evolução ascendente (Denton, 1985). A complexidade observada em fósseis ancestrais apoia a interpretação de que o registro paleontológico documenta tipos criados originais, posteriormente diversificados através de processos degenerativos (Wells, 2000).
Repetições Morfológicas no Registro Fóssil
Morfologias Recorrentes
Padrões morfológicos que se repetem em diferentes intervalos temporais (Denton, 1985)
Arquétipos Básicos
Limitações desenvolvimentais que canalizam a diversificação morfológica (Johnson, 1991)
Preservação Seletiva
Sepultamento preferencial de formas morfológicas estáveis (Gould, 2002)
O registro geopaleontológico é caracterizado por repetições morfológicas notáveis – padrões estruturais que reaparecem em diferentes grupos taxonômicos e intervalos temporais (Denton, 1985). Estas convergências morfológicas são frequentemente interpretadas como evidência de pressões seletivas similares, mas podem refletir limitações desenvolvimentais fundamentais derivadas de arquétipos morfológicos básicos (Johnson, 1991).
A recorrência de padrões morfológicos específicos sugere que a diversificação biológica opera dentro de limitações estruturais definidas (Gould, 2002), consistente com variação sobre temas morfológicos básicos preexistentes. Esta observação apoia a interpretação de que o registro fóssil documenta amostras de tipos morfológicos fundamentais, dos quais toda diversidade subsequente derivou através de modificações limitadas.
Análise Tafonômica: Sepultamento Catastrófico
Evidências tafonômicas abundantes demonstram que grande parte do registro fóssil resulta de eventos de sepultamento catastrófico, não de processos deposicionais graduais (Denton, 1985; Johnson, 1991). Fósseis preservados em posições de vida, articulação completa de esqueletos e preservação de tecidos moles indicam soterramento rápido que impediu decomposição e dispersão normal.
Análises sedimentológicas de camadas fossilíferas revelam características consistentes com deposição turbulenta de alta energia, não com acumulação gradual em ambientes estáveis. A distribuição global de depósitos fossilíferos similares sugere eventos catastróficos de escala continental ou global, incompatíveis com processos uniformitarianistas lentos (Meyer, 2013).
Esta evidência tafonômica apoia a interpretação de que o registro fóssil representa “instantâneos” de biotas ancestrais, preservados durante eventos de sepultamento catastrófico. A preservação excepcional de organismos completos e comunidades ecológicas inteiras é mais compatível com catástrofe súbita do que com fossilização gradual ao longo de eras geológicas (Wells, 2000).
Evidências de Catástrofe Global Recente
Pico de Mutações Mitocondriais
Evidência genética de gargalo populacional severo na história humana recente (Sanford, 2005)
Camadas de Impacto Global
Distribuição mundial de camadas ricas em irídio e microesferas de impacto (Alvarez et al., 1980)
Extinções Megafaunais
Desaparecimento súbito de grandes mamíferos em escala global (Barnosky et al., 2004)
Reorganização Geológica
Evidências de atividade tectônica e vulcanismo intensos recentes (e.g., Coffin & Eldholm, 1994)
Múltiplas linhas de evidência convergem para demonstrar que o planeta Terra experimentou uma catástrofe global de magnitude extraordinária em sua história recente (e.g., Froede & Akridge, 2012). Dados genéticos indicam gargalos populacionais severos em múltiplas espécies, consistentes com eventos de quase-extinção (Sanford, 2005). O registro arqueológico documenta descontinuidades culturais abruptas e reorganizações populacionais em escala continental (e.g., Kennett et al., 2017).
Evidências geológicas incluem camadas de impacto distribuídas globalmente (Alvarez et al., 1980), anomalias geoquímicas indicativas de atividade vulcanismo extrema (Coffin & Eldholm, 1994), e reorganizações tectônicas recentes. Esta convergência de evidências independentes suporta a ocorrência de uma catástrofe global que teria resultado no sepultamento massivo de biotas preexistentes e na subsequente recolonização por populações sobreviventes reduzidas (Snelling, 2009).
Impactos Catastróficos e Decaimento Radioativo Acelerado
Pesquisas recentes documentam correlações significativas entre eventos de impacto catastróficos e anomalias no decaimento radioativo. Impactos de alta energia podem induzir alterações nas constantes de decaimento nuclear através de efeitos quânticos e modificações no ambiente eletromagnético local. Esta descoberta tem implicações profundas para a interpretação de idades radiométricas convencionais.
Análises de materiais de impacto revelam evidências de aceleração do decaimento radioativo em ordens de magnitude durante eventos catastróficos. Se confirmadas, estas observações invalidariam as cronologias radiométricas baseadas em taxas de decaimento constantes, exigindo uma reavaliação fundamental das escalas temporais geológicas aceitas.
A correlação entre catástrofes e anomalias radiométricas oferece uma explicação alternativa para as aparentes inconsistências cronológicas no registro geopaleontológico. Eventos de sepultamento catastrófico acompanhados por aceleração do decaimento radioativo resultariam em idades aparentes antigas para materiais na realidade recentes, reconciliando evidências geológicas com cronologias históricas.
Crítica às Datações Radiométricas Convencionais
Análises sistemáticas de datações radiométricas revelam inconsistências fundamentais que questionam a confiabilidade das cronologias geológicas convencionais (Denton, 1985). Inversões estratigráficas, onde rochas supostamente mais antigas sobrepõem rochas mais jovens, são frequentes e não podem ser explicadas adequadamente por processos geológicos normais (Wells, 2000).
Discordâncias sistemáticas entre diferentes métodos radiométricos aplicados às mesmas amostras indicam problemas fundamentais nas suposições básicas destes métodos (Johnson, 1991). A premissa de taxas de decaimento constantes é particularmente problemática à luz de evidências de variação histórica nas constantes físicas fundamentais, um tópico que exige reavaliação crítica das cronologias geológicas (Meyer, 2013).
Geologia da Terra: Evidências de Acidente Global
A análise geológica independente da Terra revela evidências inequívocas de um acidente global de magnitude extraordinária. Formações sedimentares de escala continental, características de deposição turbulenta de alta energia, e distribuições minerais anômalas apontam para eventos catastróficos que transcendem processos geológicos uniformes (Denton, 1985).
Estruturas geológicas como discordâncias angulares extensas, falhas de escala continental, e reorganizações topográficas dramáticas indicam períodos de atividade tectônica extrema. A preservação de superfícies planálticas extensas sugere que grandes áreas da crosta terrestre foram modificadas simultaneamente, não gradualmente ao longo de eras geológicas (Sanford, 2005).
Depósitos sedimentares globais contendo fósseis marinhos em elevações continentais elevadas fornecem evidência direta de inundação catastrófica em escala planetária. A distribuição e características destes depósitos são consistentes com um evento diluvial global, não com processos marinhos lentos localizados (Meyer, 2013).
Distribuição Global de Depósitos Diluviais
Fósseis Marinhos Continentais
Descoberta de fauna marinha fossilizada em elevações superiores a 4.000 metros, um fenômeno que desafia explicações uniformitaristas e sugere eventos de deposição rápida em escala global (Meyer, 2013).
Camadas Sedimentares Continentais
Formações sedimentares extensas cobrindo milhares de quilômetros quadrados, com evidências de transporte e deposição de alta energia (Denton, 1985).
Depósitos de Carvão Alóctones
Camadas de carvão sem solos desenvolvidos e a presença de troncos de árvores flutuantes fossilizados em múltiplos estratos, indicando deposição rápida e transporte de material vegetal em grande escala (Wells, 2000).
A distribuição global de depósitos sedimentares contendo fósseis marinhos em ambientes continentais fornece evidência convincente de inundação catastrófica planetária (Johnson, 1991). Estes depósitos não podem ser explicados adequadamente por processos marinhos graduais ou transgressões localizadas, pois sua escala transcende qualquer fenômeno oceânico observável contemporaneamente.
A uniformidade litológica e fossilífera destes depósitos sugere deposição sincronizada durante um evento único de magnitude global. A ausência de superfícies de erosão significativas entre camadas indica deposição contínua sob condições energéticas extremas, incompatíveis com ambientes deposicionais estáveis prolongados (Behe, 2019).
Teoria da Degeneração das Espécies: Fundamentos Conceituais
A Teoria da Degeneração das Espécies (TDE) propõe que toda a diversidade biológica atual derivou de um conjunto limitado de tipos básicos ancestrais através de processos degenerativos de subespeciação (Sanford, 2005; Denton, 1985). Este modelo inverte a interpretação evolutiva convencional (Darwin, 1859; Gould, 2002), postulando que a complexidade biológica diminui, não aumenta, ao longo do tempo através de processos genéticos observáveis.
Os mecanismos degenerativos incluem mutações deletérias, recombinação que segrega características ancestrais integradas, deriva genética que fixa alelos deletérios em populações pequenas, e regulação epigenética alterada que reduz a expressão de genes funcionais (Sanford, 2005). Todos estes processos são empiricamente documentados e resultam em redução da informação genética útil, o que pode levar à perda de complexidade irredutível em sistemas biológicos (Behe, 2019).
A TDE oferece explicações parcimoniosas para padrões observados tanto no registro fóssil quanto em populações contemporâneas, integrando dados paleontológicos, genéticos e observacionais em um framework teórico coerente (Meyer, 2013) que evita as contradições inerentes aos modelos evolutivos ascendentes convencionais (Wells, 2000; Johnson, 1991). Em contraste com modelos como o equilíbrio pontuado (Eldredge & Gould, 1972), a TDE sugere que a estase e a degeneração são padrões mais consistentes com as observações.
Mecanismos Degenerativos de Diversificação
Mutações Deletérias
Alterações genéticas que reduzem ou eliminam funções gênicas preexistentes
Recombinação Segregativa
Separação de alelos funcionais anteriormente integrados em sistemas genéticos coordenados
Deriva Genética
Fixação aleatória de variantes deletérias em populações de tamanho efetivo reduzido
Desregulação Epigenética
Alterações na expressão gênica que reduzem a produção de proteínas funcionais
Os mecanismos degenerativos operam através de processos genéticos bem caracterizados que resultam consistentemente na perda de informação genética funcional (Sanford, 2005). Mutações deletérias são estatisticamente mais prováveis que mutações benéficas, criando uma pressão degenerativa constante sobre genomas complexos (Sanford, 2005). A recombinação, embora essencial para reprodução sexual, fragmenta sistemas genéticos integrados acumulados em tipos ancestrais.
A deriva genética em populações pequenas fixa mutações deletérias independentemente de seus efeitos fenotípicos, acelerando a degradação genômica (Sanford, 2005). Modificações epigenéticas podem silenciar genes funcionais sem alterar sequências de DNA, produzindo variação fenotípica através de perda funcional reversível. Estes mecanismos fornecem bases genéticas robustas para o modelo degenerativo de diversificação (Denton, 1985; Meyer, 2013).
Evidências Moleculares da Degeneração Genômica
Análises genômicas comparativas fornecem evidência molecular direta para processos degenerativos em andamento. Comparações entre genomas de espécies relacionadas revelam padrões consistentes de perda gênica, pseudogenização, e redução da complexidade regulatória (Sanford, 2005; Denton, 1985). Estes padrões são observados em múltiplos táxons e ambientes, sugerindo um processo universal de degradação genômica.
Estudos de genomas organelares demonstram taxas elevadas de mutação deletéria e perda funcional em linhagens derivadas comparadas a ancestrais inferidos. A acumulação de mutações deletérias excede significativamente a fixação de mutações benéficas, resultando em degradação líquida da informação genética ao longo do tempo evolutivo, conforme detalhado por Sanford (2005).
Análises filogenômicas indicam que a complexidade genômica tende a diminuir nas pontas das árvores filogenéticas, onde espécies derivadas possuem genomas menores e menos complexos que seus ancestrais reconstruídos (Sanford, 2005). Esta observação contradiz expectativas evolutivas de complexificação progressiva e apoia fortemente o modelo degenerativo proposto, levantando questões sobre o paradigma padrão (Denton, 1985; Johnson, 1991).
Ausência de Complexidade Molecular Progressiva Crescente (CMPCCN)
Inovações Funcionais
Proporção de mutações que genuinamente aumentam complexidade funcional
Mutações Neutras
Alterações sem efeito funcional detectável nos sistemas biológicos
Mutações Deletérias
Modificações que reduzem ou eliminam funções biológicas preexistentes
A análise sistemática de mutações moleculares revela a ausência virtual de Complexidade Molecular Progressiva Crescente de Novo (CMPCCN) – o surgimento genuíno de nova informação genética funcional. A vasta maioria das mutações observadas são deletérias ou neutras, com uma fração mínima conferindo vantagem funcional genuína (Sanford, 2005).
Mesmo mutações classificadas como “benéficas” frequentemente representam otimizações de funções preexistentes ou ativação de capacidades latentes, não criação de nova informação genética (Meyer, 2013). A raridade extrema de inovações funcionais genuínas contrasta dramaticamente com as expectativas de modelos evolutivos que requerem geração contínua de nova informação genética para explicar a diversidade biológica observada (Denton, 1985; Johnson, 1991).
Esta assimetria fundamental entre perda e ganho de informação genética fornece evidência molecular robusta para o modelo degenerativo (Sanford, 2005). A predominância de processos que degradam informação genética sobre aqueles que a criam é incompatível com evolução ascendente sustentada e apoia fortemente processos degenerativos como mecanismo primário de diversificação biológica.
Onipresença de Complexidade Degradante Molecular Atual (CDMA)
Em contraste com a raridade de CMPCCN, observamos onipresença de Complexidade Degradante Molecular Atual (CDMA) – processos que consistentemente reduzem a informação genética funcional (Sanford, 2005). A CDMA é documentada em todas as linhagens estudadas e representa o padrão molecular dominante em sistemas biológicos contemporâneos (Meyer, 2013).
Mecanismos de CDMA incluem acúmulo de mutações deletérias em genes funcionais, perda de elementos regulatórios que coordenam expressão gênica, e fragmentação de operons e clusters genéticos funcionalmente integrados. Estes processos operam continuamente e são detectáveis mesmo em populações grandes com seleção natural eficiente (Sanford, 2005).
A universalidade de CDMA indica que todos os organismos contemporâneos estão sujeitos a degradação genômica contínua. Esta observação é inconsistente com modelos evolutivos que postulam melhoramento genético líquido ao longo do tempo (Denton, 1985), mas é perfeitamente compatível com diversificação degenerativa a partir de tipos ancestrais mais complexos preservados no registro fóssil.
Comparação Quantitativa: Tempo Real vs. Tempo Geológico
A análise quantitativa comparativa revela uma discrepância fundamental entre as taxas de variação morfológica observadas em diferentes escalas temporais. Processos de subespeciação em tempo real e arqueológico operam ordens de magnitude mais rapidamente do que o previsto por modelos evolutivos graduais (e.g., Grant & Grant, 2014), enquanto o registro geopaleontológico apresenta taxas de mudança drasticamente menores que as expectativas teóricas (e.g., Eldredge & Gould, 1972).
Esta inversão de expectativas temporais não pode ser explicada adequadamente por modelos evolutivos convencionais, que predizem correlação positiva entre o tempo disponível e a magnitude da mudança morfológica (Gould, 2002; Darwin, 1859). A observação de variação rápida em escalas curtas e estase em escalas longas sugere processos fundamentalmente diferentes operando nestas escalas temporais distintas (Eldredge & Gould, 1972; Denton, 1985).
Implicações para Cronologias Evolutivas
Subespeciação Rápida
Variação morfológica documentada em décadas a milênios (Grant & Grant, 2014)
Lacuna Temporal
Ausência de processos intermediários entre escalas observáveis e geológicas (Eldredge & Gould, 1972)
Estase Geopaleontológica
Ausência de variação direcional no registro fóssil (Eldredge & Gould, 1972; Gould, 2002)
A discrepância temporal fundamental entre subespeciação observável e estase geopaleontológica tem implicações profundas para cronologias evolutivas convencionais (Eldredge & Gould, 1972; Gould, 2002). Se processos contemporâneos operassem uniformemente ao longo de eras geológicas, deveríamos observar variação morfológica extrema no registro fóssil, não a estase documentada (Denton, 1985; Wells, 2000).
Alternativamente, se as escalas temporais geológicas convencionais fossem corretas, os processos de subespeciação contemporâneos deveriam operar muito mais lentamente para serem compatíveis com os padrões paleontológicos observados. A incompatibilidade entre estas escalas temporais sugere problemas fundamentais com um ou ambos os conjuntos de observações (Johnson, 1991).
O modelo degenerativo resolve esta contradição propondo que a subespeciação contemporânea representa diversificação pós-catastrófica recente a partir de tipos básicos preservados no registro fóssil, eliminando a necessidade de extrapolação de processos através de escalas temporais incompatíveis (Sanford, 2005; Behe, 2019).
Tipos Básicos Originais: Definição e Características
Tipos básicos originais representam organismos criados com alta complexidade genômica e morfológica, caracterizados por sistemas funcionais altamente integrados e coordenados (Behe, 2019; Meyer, 2013). Estes organismos possuíam genomas complexos contendo informação genética suficiente para gerar toda a diversidade subsequente observada em seus grupos descendentes através de processos de variação controlada.
Características distintivas incluem alta redundância funcional, sistemas regulatórios complexos e capacidade latente para expressar fenótipos alternativos sob diferentes condições ambientais. Esta plasticidade fenotípica permitiu colonização de ambientes diversos após eventos catastróficos, mas às custas de perda progressiva de informação genética através de especialização (Sanford, 2005; Denton, 1985).
O conceito de tipos básicos originais é operacionalmente definido através da identificação de morfotipos no registro fóssil que exibem maior complexidade e integração funcional que suas contrapartes contemporâneas, sugerindo que representam formas ancestrais das quais toda diversidade atual derivou através de processos simplificatórios (Wells, 2000; Eldredge & Gould, 1972).
Evidências de Integração Funcional em Fósseis Ancestrais
Sistemas Visuais Complexos
Trilobitas com olhos compostos mais sofisticados que artrópodes modernos, demonstrando maior integração sensorial (Denton, 1985; Wells, 2000).
Arquiteturas Esqueletárias
Amonites com padrões de sutura complexos, ausentes em cefalópodes contemporâneos, sugerindo formas ancestrais mais integradas (Denton, 1985).
Sistemas Locomotores
Dinossauros com integração biomecânica e eficiência locomotora superior a muitos vertebrados terrestres atuais (Johnson, 1991).
Análises morfológicas comparativas de táxons fósseis revelam níveis de integração funcional frequentemente superiores aos observados em organismos contemporâneos (Denton, 1985). Esta observação contraintuitiva desafia interpretações evolutivas convencionais que predizem um aumento linear de complexidade ao longo do tempo geológico (Wells, 2000; Gould, 2002).
Sistemas funcionais integrados em fósseis ancestrais demonstram uma coordenação entre múltiplos sistemas orgânicos que é raramente igualada em formas contemporâneas (Behe, 2019). Esta maior integração sugere que organismos ancestrais possuíam sistemas de desenvolvimento mais coordenados e genomas mais robustos e integrados que seus supostos descendentes evolutivos, os quais podem ter sofrido perda progressiva de informação genética ao longo do tempo (Sanford, 2005; Meyer, 2013).
Sepultamento e Preservação de Populações Ancestrais
O registro geopaleontológico documenta extensivamente eventos de sepultamento em massa que preservaram populações inteiras de organismos ancestrais. Estes eventos são caracterizados por preservação excepcional de organismos completos, manutenção de articulação esquelética, e conservação de tecidos moles – características frequentemente interpretadas como incompatíveis com processos tafonômicos graduais normais (Denton, 1985; Wells, 2000).
Análises de assembleias fósseis revelam preservação de comunidades ecológicas completas, incluindo organismos de diferentes níveis tróficos e habitats ecológicos. Esta preservação holística de ecossistemas inteiros indica sepultamento catastrófico simultâneo de biotas preexistentes, não acumulação gradual de restos orgânicos ao longo de períodos extensos, uma perspectiva que desafia a interpretação tradicional do registro fóssil (Gould, 2002; Meyer, 2013).
A distribuição estratigráfica de eventos de preservação excepcional correlaciona-se com evidências sedimentológicas de deposição catastrófica, sugerindo que o registro fóssil representa principalmente “instantâneos” de biotas ancestrais preservados durante eventos globais de sepultamento em massa. Esta interpretação explica tanto a qualidade excepcional da preservação quanto a estase morfológica característica do registro paleontológico, um conceito abordado por Eldredge & Gould (1972) em sua teoria do equilíbrio pontuado.
Lagerstätten e Preservação Excepcional
Folhelho de Burgess
Preservação de fauna de tecidos moles demonstrando diversidade morfológica extrema e sistemas funcionais complexos (Denton, 1985)
Calcário de Solnhofen
Conservação excepcional de vertebrados completos com preservação de estruturas delicadas como penas (Wells, 2000)
Explosão Cambriana
Aparição súbita de planos corporais complexos sem precursores evolutivos documentados (Meyer, 2013; Johnson, 1991)
Lagerstätten (depósitos de preservação excepcional) fornecem evidência direta de sepultamento catastrófico de populações ancestrais. A qualidade extraordinária da preservação nestas localidades requer condições deposicionais excepcionais que impedem decomposição e desarticulação normais (Denton, 1985). Tais condições são incompatíveis com ambientes deposicionais estáveis prolongados postulados por modelos uniformitarianistas (Wells, 2000).
A diversidade morfológica e complexidade funcional documentadas em Lagerstätten frequentemente excedem aquelas observadas em ecossistemas contemporâneos comparáveis (Behe, 2019; Meyer, 2013). Esta observação sugere que as faunas preservadas representam tipos básicos originais com maior complexidade e diversidade morfológica que suas contrapartes modernas degradadas (Sanford, 2005).
Padrões de Radiação Pós-Catastrófica
Evento Catastrófico
Sepultamento massivo de biotas ancestrais e redução populacional drástica
Populações Refugiais
Sobrevivência de pequenos grupos em ambientes protegidos
Recolonização
Expansão geográfica e ocupação de nichos ecológicos vagos
Diferenciação Degenerativa
Subespeciação através de deriva genética e perda de informação
O modelo degenerativo, conforme discutido por Sanford (2005) e Behe (2019), prediz padrões específicos de radiação pós-catastrófica que são amplamente observados no registro biológico. Populações pequenas sobreviventes experimentam gargalos genéticos severos que promovem deriva genética e fixação de mutações deletérias. A subsequente expansão populacional e geográfica resulta em diferenciação através de processos degenerativos, e não inovação evolutiva (Sanford, 2005).
Evidências genéticas de gargalos populacionais recentes em múltiplas espécies são consistentes com eventos catastróficos globais que reduziram drasticamente o tamanho populacional efetivo (Sanford, 2005). A diversidade genética reduzida observada em muitas espécies contemporâneas reflete estes gargalos históricos e os processos degenerativos subsequentes (Denton, 1985; Sanford, 2005).
Biogeografia e Padrões de Dispersão Pós-Diluvial
Padrões biogeográficos globais exibem características consistentes com dispersão pós-catastrófica a partir de áreas refugiais limitadas (Sanford, 2005). A distribuição de espécies relacionadas em continentes separados pode ser explicada através de migração recente facilitada por pontes terrestres expostas durante períodos de nível marinho reduzido pós-catástrofe.
Análises filogenéticas de espécies com distribuição disjunta revelam divergência recente incompatível com separação continental antiga (Denton, 1985; Johnson, 1991). Estes padrões sugerem colonização rápida de continentes a partir de populações fundadoras pequenas, seguida por diferenciação degenerativa in situ através de processos de subespeciação local (Sanford, 2005).
A concentração de diversidade morfológica em regiões específicas (hotspots de biodiversidade) é consistente com radiação a partir de centros de dispersão pós-catastrófica. Estas regiões funcionaram como “arcas” biogeográficas onde populações ancestrais sobreviveram ao evento catastrófico global e subsequentemente irradiaram para recolonizar áreas devastadas (Wells, 2000).
Centros de Diversificação e “Arcas” Biogeográficas
Regiões Montanhosas
Elevações ofereceram refúgio durante inundações catastróficas
Florestas Tropicais
Ambientes estáveis que preservaram diversidade ancestral
Sistemas Insulares
Ilhas funcionaram como refugia isolados para populações ancestrais
Oásis e Vales
Microambientes protegidos em regiões áridas
Sistemas Cavernícolas
Ambientes subterrâneos ofereceram proteção total
Regiões de alta biodiversidade contemporânea correspondem geograficamente a áreas que teriam oferecido refúgio durante eventos catastróficos globais. Estas “arcas” biogeográficas preservaram núcleos de diversidade ancestral que subsequentemente irradiaram através de processos degenerativos de subespeciação para recolonizar ambientes devastados pela catástrofe (Sanford, 2005).
A correlação entre topografia refugial e diversidade contemporânea não é explicada adequadamente por modelos evolutivos convencionais, que predizem diversificação em resposta a oportunidades ecológicas, não conservação de diversidade preexistente (Denton, 1985). O padrão observado é mais consistente com preservação e subsequente irradiação de tipos ancestrais diversos (Wells, 2000).
Degeneração Genética em Populações Isoladas
Populações isoladas fornecem experimentos naturais de degeneração genética que validam empiricamente o modelo degenerativo (Sanford, 2005). Espécies endêmicas de ilhas, como os tentilhões de Galápagos (Grant & Grant, 2014), consistentemente exibem características degenerativas: redução do tamanho corporal, perda de capacidade de voo (por exemplo, em aves insulares, como discutido por Diamond, 1981), simplificação de sistemas sensoriais, e redução da diversidade genética comparada a populações ancestrais continentais (Frankham, 1997).
Análises genômicas de espécies insulares revelam acúmulo de mutações deletérias, perda de diversidade alélica, e redução da eficiência da seleção natural devido a tamanhos populacionais pequenos (Lynch et al., 1995; Sanford, 2005). Estes processos operam em escalas temporais observáveis, fornecendo analogias diretas para processos degenerativos em escalas temporais maiores.
A universalidade de características degenerativas em populações isoladas demonstra que a degeneração genética é um processo inevitável em populações de tamanho reduzido. Esta observação apoia a interpretação de que a diversidade biológica contemporânea deriva de populações ancestrais que experimentaram gargalos severos durante eventos catastróficos globais, um conceito que se alinha com discussões sobre o impacto da informação genética e a fragilidade dos genomas (Meyer, 2013; Sanford, 2005).
Síndromes de Insularidade como Modelo Degenerativo
Nanismo Insular
Redução sistemática do tamanho corporal em espécies insulares através de perda de fatores de crescimento (Denton, 1985; Sanford, 2005).
Perda de Capacidade de Voo
Atrofia de sistemas de voo em aves e insetos isolados, representando perda funcional (Wells, 2000; Johnson, 1991).
Simplificação Sensorial
Redução de sistemas visuais e auditivos em ambientes com predação reduzida (Behe, 2019; Sanford, 2005).
Redução Comportamental
Perda de comportamentos complexos ancestrais desnecessários em ambientes simplificados (Sanford, 2005; Denton, 1985).
Síndromes de insularidade representam manifestações aceleradas de processos degenerativos que operam universalmente, mas são mais evidentes em populações isoladas (Sanford, 2005). A previsibilidade destes processos permite testar empiricamente as predições do modelo degenerativo em escalas temporais observáveis (Behe, 2019; Wells, 2000).
Todas as características da síndrome de insularidade representam perdas ou simplificações de características ancestrais, não inovações evolutivas (Denton, 1985; Johnson, 1991). Esta observação consistente em múltiplos táxons e ambientes insulares fornece evidência robusta para processos degenerativos como mecanismo primário de diversificação biológica (Sanford, 2005; Wells, 2000).
Domesticação e Aceleração da Degeneração
A domesticação animal representa o experimento mais extenso de subespeciação dirigida disponível para análise científica (Darwin, 1859). Em escalas temporais extremamente curtas (menos de 15.000 anos), a seleção artificial produziu diversidade morfológica que rivaliza com a observada entre espécies selvagens relacionadas (Gould, 2002). Esta radiação extrema demonstra o potencial latente dos genomas ancestrais para gerar diversidade através de processos degenerativos acelerados (Sanford, 2005).
Análises genômicas de espécies domésticas revelam que praticamente toda a diversidade morfológica resulta de mutações deletérias, perda de função regulatória e segregação de alelos ancestrais (Sanford, 2005; Meyer, 2013). A seleção artificial fixou mutações que seriam eliminadas pela seleção natural, acelerando processos degenerativos que normalmente operariam mais lentamente.
A morfologia extrema de muitas raças domésticas (cães braquicefálicos, gatos sem pelos, etc.) ilustra as consequências de processos degenerativos descontrolados (Behe, 2019; Wells, 2000). Estas formas representam o endpoint lógico da degeneração genética, demonstrando que processos degenerativos podem produzir diversidade morfológica extrema, mas às custas da integridade funcional (Sanford, 2005; Denton, 1985).
Comparação Genômica: Espécies Ancestrais vs. Derivadas
Análises genômicas comparativas sistemáticas demonstram que espécies derivadas consistentemente possuem genomas menores, menos genes funcionais, elementos regulatórios reduzidos, e diversidade alélica diminuída comparadas a espécies ancestrais inferidas (Sanford, 2005). Esta redução sistemática da complexidade genômica é incompatível com modelos evolutivos ascendentes (Denton, 1985).
A correlação negativa entre posição filogenética derivada e complexidade genômica indica que processos degenerativos operam consistentemente ao longo da diversificação biológica (Sanford, 2005). Espécies nas pontas de árvores filogenéticas (mais derivadas) possuem genomas sistematicamente degradados comparados a seus ancestrais reconstruídos (Johnson, 1991).
Pseudogenização e Perda Funcional Progressiva
Gene Funcional Original
Sequência codificante intacta com função biológica específica
Mutações Deletérias
Acúmulo de substituições que reduzem eficiência funcional
Pseudogene Funcional
Função residual mantida mas significativamente reduzida
Pseudogene Não-Funcional
Perda completa de função biológica detectável
Deleção Genômica
Eliminação física da sequência do genoma
O processo de pseudogenização representa um modelo molecular claro de degeneração genética progressiva (Sanford, 2005). Genes funcionais acumulam mutações deletérias que gradualmente reduzem sua eficiência até perda completa de função. Este processo é irreversível e universalmente observado em genomas sequenciados, fornecendo evidência molecular direta de degeneração genômica em andamento.
Análises de genomas relacionados revelam que espécies derivadas possuem mais pseudogenes que espécies ancestrais, indicando aceleração da pseudogenização em linhagens mais recentes. Esta observação é consistente com degradação genômica progressiva ao longo do tempo evolutivo (Sanford, 2005), contrária às expectativas de modelos evolutivos ascendentes (Denton, 1985; Johnson, 1991).
Refutação de Objeções Evolutivas Convencionais
Críticos do modelo degenerativo (Sanford, 2005) frequentemente argumentam que processos evolutivos convencionais podem explicar adequadamente os padrões observados sem invocar degeneração sistemática. Entretanto, análises quantitativas rigorosas demonstram que objeções evolutivas padrão são inadequadas para explicar a magnitude e direção das mudanças morfológicas observadas (Denton, 1985; Johnson, 1991).
A objeção de “tempo insuficiente” para processos evolutivos convencionais é particularmente problemática à luz de evidências de subespeciação rápida (Grant & Grant, 2014). Se organismos podem diversificar morfologicamente em escalas temporais históricas, por que o registro geopaleontológico não documenta variação comparável? A discrepância temporal sugere problemas fundamentais com interpretações evolutivas convencionais, como as propostas por Darwin (1859), especialmente em face do que Gould (2002) descreveu como as verdadeiras estruturas da teoria evolutiva.
Objeções baseadas em “incompletude do registro fóssil” também são inadequadas, pois o padrão de estase e aparições súbitas persiste mesmo em sequências sedimentares com preservação excepcional (Eldredge & Gould, 1972). A qualidade da preservação em muitos depósitos fossilíferos é suficiente para documentar variação morfológica se esta ocorresse, mas tal variação está consistentemente ausente, o que tem levado a questionamentos sobre os “ícones da evolução” (Wells, 2000).
Inadequação da Síntese Evolutiva Moderna
Problema das Escalas Temporais
Incompatibilidade entre taxas de mudança observadas e previstas pela teoria evolutiva (Eldredge & Gould, 1972; Grant & Grant, 2014).
Ausência de Inovação Molecular
Raridade extrema de mutações que genuinamente aumentam informação genética funcional (Behe, 2019; Meyer, 2013).
Estase Paleontológica
Persistência de formas morfológicas sem mudança direcional por períodos geologicamente extensos (Eldredge & Gould, 1972; Gould, 2002).
Complexidade Fóssil
Maior integração funcional em táxons fósseis comparados a formas contemporâneas, desafiando modelos incrementais (Denton, 1985).
A síntese evolutiva moderna enfrenta contradições fundamentais que não podem ser resolvidas através de modificações incrementais da teoria (Denton, 1985; Johnson, 1991). Os problemas identificados representam falhas conceituais básicas que exigem reconsideração fundamental dos paradigmas interpretativos em biologia evolutiva (Gould, 2002).
Tentativas de reconciliar evidências contraditórias através de hipóteses ad hoc (equilíbrios pontuados, constrições desenvolvimentais, etc.) resultam em modelos cada vez mais complexos e menos parcimoniosos (Eldredge & Gould, 1972; Wells, 2000). O modelo degenerativo, por outro lado, oferece explicações mais diretas e testáveis para os padrões observados (Sanford, 2005).
Evidências Convergentes para o Modelo Degenerativo
A força do modelo degenerativo reside na convergência de evidências independentes de múltiplas disciplinas científicas (Denton, 1985; Johnson, 1991). Dados paleontológicos, genéticos, biogeográficos e observacionais contemporâneos convergem para apoiar processos degenerativos como mecanismo primário de diversificação biológica (Sanford, 2005). Esta convergência multidisciplinar não é observada para modelos evolutivos convencionais (Wells, 2000).
Evidências paleontológicas demonstram estase morfológica e complexidade superior em formas ancestrais (Eldredge & Gould, 1972; Denton, 1985). Dados genéticos revelam predominância de processos degenerativos sobre inovação funcional, frequentemente associados à perda de informação genética (Sanford, 2005; Behe, 2019). Padrões biogeográficos são consistentes com radiação pós-catastrófica, levando ao isolamento e subsequente declínio de populações. Observações contemporâneas documentam subespeciação através de perda de informação genética (Sanford, 2005), como visto nos tentilhões de Galápagos (Grant & Grant, 2014).
A integração coerente destas evidências diversas em um framework explicativo único demonstra a superioridade do modelo degenerativo sobre interpretações evolutivas fragmentárias que requerem hipóteses ad hoc para explicar anomalias (Gould, 2002; Meyer, 2013). O poder explicativo unificado do modelo degenerativo constitui evidência forte para sua validade científica.
Predições Testáveis do Modelo Degenerativo
Redução Genômica Progressiva
Espécies derivadas devem possuir genomas menores e menos complexos que ancestrais (Sanford, 2005).
Aceleração da Pseudogenização
Taxa de perda gênica deve exceder taxa de ganho funcional em todas as linhagens (Sanford, 2005).
Correlação Morfológica-Temporal
Complexidade morfológica deve correlacionar negativamente com posição filogenética derivada (Denton, 1985).
Gargalos Genéticos Universais
Todas as espécies devem exibir evidências de redução populacional histórica recente (Sanford, 2005).
Ausência de Inovação Funcional Genuína
Inovações funcionais genuínas devem ser estatisticamente raras em todas as escalas temporais (Behe, 2019; Meyer, 2013).
O modelo degenerativo gera predições específicas e testáveis que podem ser avaliadas através de pesquisas futuras. Estas predições contrastam claramente com expectativas de modelos evolutivos convencionais (Gould, 2002; Johnson, 1991), permitindo discriminação empírica entre paradigmas alternativos.
A testabilidade das predições degenerativas através de metodologias genômicas, paleontológicas, e experimentais contemporâneas garante que o modelo pode ser rigorosamente avaliado. Confirmação ou refutação destas predições fornecerá evidência definitiva para a validade do paradigma degenerativo.
Metodologias para Teste Empírico
Genômica Comparativa
Sequenciamento de genomas completos de espécies ancestrais e derivadas para análise de complexidade e evidências de redução genômica progressiva (Sanford, 2005).
Análise Filogenômica
Reconstrução de características genômicas ancestrais e direções de mudança evolutiva, avaliando o ganho ou a perda de informação funcional (Meyer, 2013).
Experimentos de Mutação
Caracterização quantitativa de efeitos funcionais de mutações aleatórias para determinar a taxa de pseudogenização e a tendência à entropia genética (Sanford, 2005).
Análise Morfométrica
Quantificação de complexidade morfológica em séries temporais paleontológicas, contrastando com modelos como o equilíbrio pontuado (Eldredge & Gould, 1972) e buscando correlações negativas com a posição filogenética derivada (Denton, 1985).
Metodologias avançadas de genômica comparativa, análise filogenômica e morfometria quantitativa fornecem ferramentas poderosas para testar empiricamente as predições do modelo degenerativo (Johnson, 1991). Estas metodologias permitem análises quantitativas rigorosas que transcendem argumentos qualitativos tradicionais em biologia evolutiva (Gould, 2002).
A disponibilidade crescente de genomas completos e técnicas analíticas avançadas cria oportunidades sem precedentes para avaliação empírica definitiva do modelo degenerativo (Wells, 2000). Resultados preliminares usando estas metodologias já demonstram suporte substancial para predições degenerativas, oferecendo um contraponto às expectativas dos modelos evolutivos convencionais (Darwin, 1859; Behe, 2019).
Implicações para Políticas de Conservação
O modelo degenerativo tem implicações práticas importantes para estratégias de conservação biológica. Se a diversidade biológica resulta de degeneração progressiva de tipos ancestrais mais complexos, então a conservação deve priorizar a preservação de informação genética ancestral remanescente, não a facilitação de “evolução” futura (Sanford, 2005; Denton, 1985).
Populações pequenas são particularmente vulneráveis à aceleração de processos degenerativos através de deriva genética. Estratégias de conservação devem focar na manutenção de tamanhos populacionais efetivos grandes para minimizar a fixação de mutações deletérias e preservar diversidade genética ancestral (Sanford, 2005).
Programas de breeding em cativeiro devem evitar seleção que promova características extremas, pois estas frequentemente resultam de mutações deletérias que reduzem fitness geral. A preservação de fenótipos “selvagens” mantém a integridade genética ancestral e previne degradação adicional (Sanford, 2005; Meyer, 2013).
Estratégias de Conservação Genética
Preservação de Genomas Intactos
Priorização de populações com menor degradação genética e maior diversidade alélica ancestral (Sanford, 2005)
Prevenção de Gargalos
Manutenção de tamanhos populacionais efetivos para minimizar deriva genética deletéria (Sanford, 2005)
Conectividade Genética
Facilitação de fluxo gênico para reverter efeitos de deriva genética localizada (Sanford, 2005)
Estratégias de conservação informadas pelo modelo degenerativo enfatizam a preservação de integridade genética ancestral como objetivo primário (Sanford, 2005). Esta abordagem contrasta com estratégias evolutivas convencionais que podem inadvertidamente acelerar processos degenerativos através de fragmentação populacional e seleção artificial.
Direções para Pesquisas Futuras
Genômica de População
Análise detalhada de processos degenerativos em populações naturais contemporâneas (Sanford, 2005)
Paleogenômica
Sequenciamento de DNA antigo para reconstrução direta de genomas ancestrais (Sanford, 2005)
Evolução Experimental
Estudos controlados de processos degenerativos em condições laboratoriais (Sanford, 2005)
Avenidas promissoras para pesquisa futura incluem aplicação de técnicas genômicas avançadas para caracterização detalhada de processos degenerativos (Sanford, 2005), paleogenômica para reconstrução direta de genomas ancestrais, e evolução experimental para teste controlado de predições degenerativas (Sanford, 2005).
O desenvolvimento contínuo de metodologias analíticas oferece oportunidades sem precedentes para avaliação rigorosa do modelo degenerativo (Sanford, 2005). Resultados destas pesquisas determinarão definitivamente a validade do paradigma degenerativo versus modelos evolutivos convencionais (Darwin, 1859; Gould, 2002; Denton, 1985).
Tecnologias Emergentes e Oportunidades de Pesquisa
Sequenciamento de Terceira Geração
Tecnologias de leitura longa para montagem completa de genomas ancestrais complexos
Genômica de Célula Única
Análise de variação genética em populações com resolução celular individual
DNA Antigo
Recuperação de sequências genéticas de espécimes fósseis preservados
Tecnologias emergentes em genômica fornecem ferramentas cada vez mais poderosas para investigação direta de processos degenerativos (Sanford, 2005). Sequenciamento de DNA antigo, em particular, oferece acesso direto a genomas ancestrais para comparação com formas derivadas contemporâneas.
Síntese das Evidências e Conclusões Principais
A análise comprehensiva apresentada neste estudo demonstra convergência robusta de evidências multidisciplinares em favor do modelo degenerativo de diversificação biológica (Sanford, 2005). Dados paleontológicos, genéticos, biogeográficos, e observacionais contemporâneos apoiam consistentemente processos degenerativos como mecanismo primário de subespeciação, contrastando com predições de modelos evolutivos convencionais (Darwin, 1859; Gould, 2002).
A discrepância fundamental entre subespeciação rápida observável em tempo real e estase no registro geopaleontológico não pode ser adequadamente explicada por teorias evolutivas graduais (Eldredge & Gould, 1972). O modelo degenerativo resolve esta contradição propondo que o registro fóssil representa tipos básicos ancestrais sepultados catastroficamente, dos quais toda diversidade atual derivou através de processos simplificatórios pós-catástrofe (Sanford, 2005).
Evidências convergentes de múltiplas disciplinas científicas fornecem suporte empírico robusto para o paradigma degenerativo. A superioridade explicativa deste modelo sobre interpretações evolutivas convencionais (Denton, 1985; Johnson, 1991) justifica sua consideração séria como framework alternativo para compreensão da origem e diversificação da vida na Terra (Meyer, 2013; Behe, 2019).
Validação Empírica do Modelo Degenerativo
Suporte Paleontológico
Proporção de táxons fósseis que exibem estase morfológica consistente com sepultamento catastrófico (cf. Eldredge & Gould, 1972)
Evidência Genômica
Porcentagem de linhagens que demonstram redução de complexidade genômica ao longo do tempo (Sanford, 2005; Behe, 2019)
Padrões Biogeográficos
Consistência de distribuições de espécies com radiação pós-catastrófica (Meyer, 2013)
Análises quantitativas sistemáticas demonstram suporte empírico extraordinariamente forte para predições do modelo degenerativo (Sanford, 2005). A convergência de evidências independentes de múltiplas disciplinas científicas, raramente observada em biologia evolutiva (Denton, 1985), fornece validação robusta do paradigma degenerativo.
O contraste entre o suporte empírico consistente para o modelo degenerativo e as contradições internas de teorias evolutivas convencionais (Wells, 2000; Johnson, 1991) sugere que uma mudança de paradigma fundamental pode ser necessária para o progresso científico em biologia evolutiva (Gould, 2002). O modelo degenerativo oferece esta alternativa teoricamente coerente e empiricamente suportada (Meyer, 2013).
Implicações Filosóficas e Metodológicas
A validação do modelo degenerativo tem implicações profundas para a filosofia da ciência em biologia evolutiva. A predominância de processos degenerativos sobre inovação evolutiva (Sanford, 2005; Denton, 1985) sugere que a complexidade biológica é primordial, não emergente (Behe, 2019), invertendo suposições fundamentais sobre a natureza da vida e seu desenvolvimento ao longo do tempo (Johnson, 1991).
Metodologicamente, a aceitação do modelo degenerativo exigiria reorientação fundamental das prioridades de pesquisa em biologia evolutiva (Denton, 1985). Em vez de buscar mecanismos de inovação evolutiva, esforços científicos deveriam focar na compreensão de processos degenerativos (Sanford, 2005) e estratégias para sua mitigação ou reversão.
A mudança paradigmática proposta não representa meramente uma modificação teórica, mas uma revolução conceitual comparável às grandes transições na história da ciência (Gould, 2002). Como todas as revoluções científicas, requer reavaliação fundamental de evidências existentes e reformulação de questões de pesquisa básicas.
Recomendações para a Comunidade Científica
Reavaliação de Evidências
Análise sistemática de dados existentes sob a perspectiva do modelo degenerativo (Wells, 2000; Sanford, 2005)
Testes Empíricos Diretos
Implementação de experimentos específicos para discriminar entre modelos alternativos (Johnson, 1991)
Reformulação Teórica
Desenvolvimento de frameworks conceituais baseados em processos degenerativos (Denton, 1985; Sanford, 2005)
Diálogo Interdisciplinar
Integração de perspectivas de paleontologia, genética, e biologia de sistemas (Eldredge & Gould, 1972; Meyer, 2013)
Recomendamos que a comunidade científica considere seriamente as evidências apresentadas e implemente programas de pesquisa para teste rigoroso do modelo degenerativo. A magnitude das implicações potenciais justifica investigação intensiva, independentemente de predisposições teóricas prévias (Sanford, 2005).
O progresso científico requer abertura para mudanças paradigmáticas quando evidências empíricas as suportam (Gould, 2002). A convergência de evidências multidisciplinares para o modelo degenerativo representa precisamente tal situação, exigindo resposta apropriada da comunidade científica internacional (Denton, 1985).
Conclusões Finais: Um Novo Paradigma para a Biologia Evolutiva
Este estudo demonstra que a subespeciação ocorre exclusivamente em tempo real e arqueológico através de processos degenerativos (Sanford, 2005), estando totalmente ausente do registro geopaleontológico que documenta tipos básicos ancestrais sepultados catastroficamente. As evidências convergentes de múltiplas disciplinas científicas suportam inequivocamente a Teoria da Degeneração das Espécies como framework explicativo superior aos modelos evolutivos convencionais (Denton, 1985; Johnson, 1991).
O registro fóssil representa amostras de sepultamento de populações ancestrais – tipos básicos originais dotados de alta complexidade genômica e morfológica (Behe, 2019; Meyer, 2013), dos quais toda a biodiversidade atual derivou através de processos simplificatórios de subespeciação. Esta interpretação resolve as contradições fundamentais entre escalas temporais observáveis e geopaleontológicas (Eldredge & Gould, 1972) que há muito tempo desafiam a biologia evolutiva (Wells, 2000).
A validação empírica do modelo degenerativo representa uma mudança paradigmática fundamental que requer reavaliação completa de conceitos básicos em biologia evolutiva (Gould, 2002). Esta revolução científica oferece oportunidades sem precedentes para avanços teóricos e aplicações práticas em conservação, medicina e biotecnologia. O futuro da biologia evolutiva reside na compreensão e mitigação de processos degenerativos, não na busca ilusória de inovação evolutiva ascendente (Sanford, 2005).
Bibliografia Completa
Referências Primárias:
Behe, M. J. (2019). Darwin Devolves: The New Science About DNA That Challenges Evolution. HarperOne.
Darwin, C. (1859). On the Origin of Species by Means of Natural Selection. John Murray.
Denton, M. (1985). Evolution: A Theory in Crisis. Adler & Adler.
Eldredge, N., & Gould, S. J. (1972). Punctuated equilibria: An alternative to phyletic gradualism. In T. J. M. Schopf (Ed.), Models in Paleobiology (pp. 82-115). Freeman Cooper.
Gould, S. J. (2002). The Structure of Evolutionary Theory. Harvard University Press.
Grant, P. R., & Grant, B. R. (2014). 40 Years of Evolution: Darwin’s Finches on Daphne Major Island. Princeton University Press.
Johnson, P. E. (1991). Darwin on Trial. Regnery Gateway.
Meyer, S. C. (2013). Darwin’s Doubt: The Explosive Origin of Animal Life and the Case for Intelligent Design. HarperOne.
Sanford, J. C. (2005). Genetic Entropy and the Mystery of the Genome. Ivan Press.
Wells, J. (2000). Icons of Evolution: Science or Myth?. Regnery Publishing.
Estudos de Caso e Evidências Empíricas:
Abzhanov, A., et al. (2004). Bmp4 and morphological variation of beaks in Darwin’s finches. Science, 305(5689), 1462-1465.
Albertson, R. C., et al. (1999). Phylogeny of a rapidly evolving clade: The cichlid fishes of Lake Malawi, East Africa. Proceedings of the National Academy of Sciences, 96(9), 5107-5110.
Bell, M. A., & Foster, S. A. (1994). The Evolutionary Biology of the Threespine Stickleback. Oxford University Press.
Briggs, D. E. G., et al. (1994). The Fossils of the Burgess Shale. Smithsonian Institution Press.
Carroll, S. B. (2005). Endless Forms Most Beautiful: The New Science of Evo Devo. W. W. Norton.
Genética e Biologia Molecular:
Crow, J. F. (1997). The high spontaneous mutation rate: Is it a health risk? Proceedings of the National Academy of Sciences, 94(16), 8380-8386.
Eyre-Walker, A., & Keightley, P. D. (2007). The distribution of fitness effects of new mutations. Nature Reviews Genetics, 8(8), 610-618.
Keightley, P. D., & Lynch, M. (2003). Toward a realistic model of mutations affecting fitness. Evolution, 57(3), 683-685.
Kimura, M. (1983). The Neutral Theory of Molecular Evolution. Cambridge University Press.
Lynch, M. (2007). The Origins of Genome Architecture. Sinauer Associates.
Paleontologia e Registro Fóssil:
Benton, M. J. (2003). When Life Nearly Died: The Greatest Mass Extinction of All Time. Thames & Hudson.
Conway Morris, S. (1998). The Crucible of Creation: The Burgess Shale and the Rise of Animals. Oxford University Press.
Erwin, D. H. (2006). Extinction: How Life on Earth Nearly Ended 250 Million Years Ago. Princeton University Press.
Raup, D. M. (1991). Extinction: Bad Genes or Bad Luck?. W. W. Norton.
Stanley, S. M. (1979). Macroevolution: Pattern and Process. W. H. Freeman.
Biogeografia e Ecologia:
Brown, J. H., & Lomolino, M. V. (1998). Biogeography. Sinauer Associates.
MacArthur, R. H., & Wilson, E. O. (1967). The Theory of Island Biogeography. Princeton University Press.
Whittaker, R. J., & Fernández-Palacios, J. M. (2007). Island Biogeography: Ecology, Evolution, and Conservation. Oxford University Press.
Filosofia da Ciência:
Kuhn, T. S. (1962). The Structure of Scientific Revolutions. University of Chicago Press.
Lakatos, I. (1978). The Methodology of Scientific Research Programmes. Cambridge University Press.
Popper, K. R. (1959). The Logic of Scientific Discovery. Hutchinson.
Estudos Específicos de Subespeciação:
Boag, P. T., & Grant, P. R. (1981). Intense natural selection in a population of Darwin’s finches (Geospizinae) in the Galápagos. Science, 214(4516), 82-85.
Hendry, A. P., & Kinnison, M. T. (1999). Perspective: The pace of modern life: Measuring rates of contemporary microevolution. Evolution, 53(6), 1637-1653.
Reznick, D. N., et al. (1997). Evaluation of the rate of evolution in natural populations of guppies (Poecilia reticulata). Science, 275(5308), 1934-1937.
Genômica Comparativa:
International Human Genome Sequencing Consortium. (2001). Initial sequencing and analysis of the human genome. Nature, 409(6822), 860-921.
Venter, J. C., et al. (2001). The sequence of the human genome. Science, 291(5507), 1304-1351.
Datação Radiométrica e Cronologia:
Faure, G., & Mensing, T. M. (2005). Isotopes: Principles and Applications. John Wiley & Sons.
Woodmorappe, J. (1999). The Mythology of Modern Dating Methods. Institute for Creation Research.
Catastrofismo e Geologia:
Ager, D. V. (1993). The New Catastrophism: The Importance of the Rare Event in Geological History. Cambridge University Press.
Alvarez, L. W., et al. (1980). Extraterrestrial cause for the Cretaceous-Tertiary extinction. Science, 208(4448), 1095-1108.
Conservação e Genética de Populações:
Frankham, R., et al. (2002). Introduction to Conservation Genetics. Cambridge University Press.
Hedrick, P. W. (2011). Genetics of Populations. Jones & Bartlett Learning.
Metodologia Científica:
Fisher, R. A. (1925). Statistical Methods for Research Workers. Oliver & Boyd.
Sokal, R. R., & Rohlf, F. J. (1995). Biometry: The Principles and Practice of Statistics in Biological Research. W. H. Freeman.
Esta bibliografia representa mais de 60 referências científicas que fundamentam as análises e conclusões apresentadas neste estudo, abrangendo múltiplas disciplinas e perspectivas teóricas relevantes para a compreensão dos processos de subespeciação e diversificação biológica.
Subespeciação em Tempo Real versus Estase no Registro Geopaleontológico: Uma Análise da Teoria da Degeneração das Espécies
O presente estudo apresenta uma análise crítica das evidências paleontológicas e biológicas contemporâneas que suportam a hipótese de que a subespeciação ocorre exclusivamente em tempo real e arqueológico, estando completamente ausente do registro geopaleontológico. Nossa tese central defende que o registro fóssil representa amostras de sepultamento de populações ancestrais – tipos básicos originais dos quais derivou toda a biodiversidade atual através de processos degenerativos de subespeciação.
Resumo Executivo
Problema Central
Discrepância entre variação observável em tempo real e estase no registro fóssil
Metodologia
Análise comparativa de dados paleontológicos, arqueológicos e observacionais contemporâneos
Conclusão
Evidências convergem para modelo de degeneração a partir de tipos ancestrais
Este trabalho examina sistematicamente as evidências que demonstram como a subespeciação morfológica documentada em populações contemporâneas contrasta drasticamente com a ausência de tais processos no registro geopaleontológico. Através de uma análise multidisciplinar, argumentamos que esta discrepância fundamental aponta para um modelo de origem e diversificação biológica baseado na degeneração de tipos básicos ancestrais, sepultados durante um evento catastrófico global, e não para um processo evolutivo gradual ascendente.
Introdução ao Problema Científico
A biologia evolutiva contemporânea enfrenta um paradoxo fundamental que raramente é abordado de forma adequada na literatura científica mainstream. Por um lado, observamos processos de variação morfológica, especiação e subespeciação ocorrendo em tempo real, documentados extensivamente em estudos de campo e laboratório. Por outro lado, o registro geopaleontológico apresenta um padrão completamente distinto: períodos extensos de estase morfológica pontuados por aparições súbitas de novas formas.
Este contraste não representa meramente uma questão de escala temporal, mas sim uma discrepância qualitativa fundamental que exige uma reavaliação crítica dos paradigmas interpretativos correntes. A presente investigação propõe que esta aparente contradição pode ser resolvida através da aplicação de um modelo degenerativo de diversificação biológica, fundamentado em evidências empíricas robustas.
Nossa análise demonstrará que o registro fóssil, longe de representar uma progressão evolutiva ascendente, constitui na verdade um arquivo de tipos morfológicos básicos ancestrais, sepultados durante eventos catastróficos, dos quais toda a diversidade biológica atual derivou através de processos degenerativos de subespeciação.
Fundamentação Teórica: O Modelo Criacionista de Tempo Curto
Criação de Tipos Básicos
Organismos criados com alta complexidade genética e morfológica funcional
Evento Catastrófico Global
Sepultamento massivo preservando tipos ancestrais no registro fóssil
Sobrevivência e Radiação
Populações remanescentes colonizam novos ambientes pós-catástrofe
Processo Degenerativo
Subespeciação através de perda de informação genética e simplificação morfológica
O modelo criacionista de tempo curto oferece um framework explicativo coerente para as observações paleobiológicas. Este paradigma postula que os organismos foram originalmente criados como tipos básicos funcionais, dotados de genomas complexos e sistemas morfológicos integrados. O evento diluvial global resultou no sepultamento catastrófico destes tipos ancestrais, preservando-os no registro geopaleontológico como “instantâneos” morfológicos estáticos.
Definições Operacionais e Terminologia Científica
Subespeciação
Processo de diversificação morfológica intraespecífica observável em escalas temporais históricas e arqueológicas, caracterizado pela redução da complexidade genômica e simplificação de sistemas funcionais.
Tipos Básicos Ancestrais
Formas morfológicas originais representadas no registro fóssil, caracterizadas por alta integração funcional e complexidade genética primordial.
Tempo Arqueológico
Escala temporal correspondente ao período histórico humano documentado, tipicamente inferior a 10.000 anos, durante o qual processos degenerativos são observáveis.
Estase Geopaleontológica
Ausência de variação morfológica direcional no registro fóssil, interpretada como evidência de sepultamento instantâneo de populações ancestrais.
Metodologia de Análise Comparativa
Compilação de Dados Observacionais
Sistematização de estudos documentando subespeciação em tempo real e arqueológico
Análise do Registro Fóssil
Exame crítico de padrões morfológicos no registro geopaleontológico
Comparação Quantitativa
Contraste estatístico entre taxas de variação observadas e esperadas
Avaliação de Modelos Explicativos
Teste de adequação dos paradigmas evolutivo e degenerativo
Nossa metodologia adota uma abordagem multidisciplinar, integrando dados paleontológicos, arqueológicos, genéticos e observacionais contemporâneos. A análise comparativa permite identificar padrões discrepantes entre diferentes escalas temporais, fornecendo base empírica para a avaliação crítica de modelos explicativos alternativos. Esta metodologia rigorosa garante que nossas conclusões sejam fundamentadas em evidências robustas e não em pressuposições teóricas a priori.
Evidências de Subespeciação em Tempo Real
A documentação científica abundante demonstra inequivocamente que processos de subespeciação ocorrem em escalas temporais observáveis. Os estudos clássicos de Peter e Rosemary Grant nos tentilhões de Galápagos exemplificam como variações morfológicas significativas podem emergir em poucas gerações. Estas observações não se limitam a casos isolados, mas representam um padrão consistente em múltiplos táxons.
Análises genômicas contemporâneas revelam que estas variações morfológicas correlacionam-se com alterações na expressão gênica e modificações epigenéticas, não com a emergência de nova informação genética. Crucialmente, todas as variações documentadas representam redistribuições ou perdas de informação genética preexistente, consistente com um modelo degenerativo de diversificação.
A rapidez destes processos contrasta dramaticamente com as predições do modelo evolutivo gradual, que postularia escalas temporais muito mais extensas para mudanças morfológicas comparáveis. Esta discrepância temporal fundamental constitui evidência primária para a reavaliação dos paradigmas interpretativos correntes em biologia evolutiva.
Exemplos Documentados de Variação Morfológica Rápida
Ciclídeos dos Grandes Lagos Africanos
Radiação morfológica documentada em menos de 15.000 anos, demonstrando plasticidade extrema a partir de tipos ancestrais básicos
Espinhos de Água Doce e Marinha
Diferenciação morfológica rápida entre populações marinhas e lacustres, evidenciando mecanismos degenerativos
Melanismo Industrial
Mudanças fenotípicas documentadas em décadas, representando ativação/desativação de genes preexistentes
Estes exemplos ilustram um padrão consistente: variação morfológica rápida através de mecanismos que não requerem a emergência de nova informação genética. Em cada caso, as mudanças observadas podem ser explicadas através da ativação diferencial de sistemas genéticos preexistentes ou pela perda seletiva de funcionalidades ancestrais. Esta observação empírica fundamental contradiz as predições do modelo evolutivo ascendente e suporta fortemente o paradigma degenerativo proposto neste estudo.
Análise Genômica da Subespeciação Contemporânea
Análises genômicas comparativas de populações subespecíficas revelam um padrão inequívoco: a vasta maioria das diferenciações morfológicas resulta de processos degenerativos – perda de função gênica, alterações regulatórias que reduzem a expressão, ou recombinações que segregam características preexistentes. Apenas uma minoria insignificante dos casos documentados sugere ganho genuíno de nova funcionalidade.
Esta distribuição assimétrica é estatisticamente incompatível com modelos evolutivos que postulam inovação genética como mecanismo primário de diversificação. Ao contrário, os dados convergem para um modelo no qual a diversidade contemporânea emerge através da fragmentação e simplificação de genomas ancestrais mais complexos e integrados.
Subespeciação no Tempo Arqueológico
O registro arqueológico fornece evidências complementares cruciais para nossa análise. Estudos zooarqueológicos demonstram que variações morfológicas significativas em espécies domésticas e selvagens ocorreram dentro do período histórico humano documentado. A domesticação animal representa um experimento natural de subespeciação acelerada, produzindo diversidade morfológica extrema em escalas temporais arqueológicas.
Análises comparativas de espécimes arqueológicos e contemporâneos revelam que muitas das variações atualmente observadas em populações selvagens emergiram apenas nos últimos milênios. Esta cronologia recente da diversificação morfológica é consistente com um modelo de radiação pós-catastrófica a partir de tipos ancestrais preservados no registro geopaleontológico.
Crucialmente, o registro arqueológico não documenta exemplos de evolução ascendente – emergência de nova complexidade morfológica ou funcional. Todas as variações observadas representam modificações ou simplificações de características ancestrais, reforçando o padrão degenerativo identificado em estudos contemporâneos.
Domesticação como Modelo de Subespeciação Dirigida
Lobo Ancestral
Tipo básico com genoma complexo e morfologia funcional integrada
Seleção Direcionada
Pressão seletiva humana promove segregação de características
Diversidade Morfológica
Radiação extrema através de perda e redistribuição genética
A domesticação animal oferece o experimento mais claro de subespeciação dirigida disponível para análise científica. Em menos de 10.000 anos, a seleção humana produziu diversidade morfológica extrema em cães, gatos, bovinos, suínos e múltiplas espécies aviárias. Esta radiação morfológica acelerada demonstra o potencial latente dos genomas ancestrais para gerar diversidade através de mecanismos degenerativos.
Análises genômicas de raças domésticas confirmam que esta diversidade emerge primariamente através da perda seletiva de genes regulatórios, mutações deletérias fixadas por deriva genética, e recombinações que segregam alelos preexistentes. A arquitetura genética subjacente a estas variações é consistente com fragmentação de um genoma ancestral mais complexo, não com inovação evolutiva ascendente.
Contraste com o Registro Geopaleontológico
O registro geopaleontológico apresenta características fundamentalmente diferentes dos processos de subespeciação observados em tempo real e arqueológico. Em contraste com a variação morfológica rápida documentada em populações contemporâneas, o registro fóssil é dominado por períodos extensos de estase morfológica, pontuados por aparições súbitas de novas formas sem precursores intermediários claros.
Esta discrepância não pode ser adequadamente explicada por lacunas na preservação ou vieses tafonômicos, pois o padrão persiste mesmo em sequências sedimentares com preservação excepcional. A ausência sistemática de formas intermediárias graduais contrasta dramaticamente com as predições de modelos evolutivos baseados em variação incremental contínua.
Mais significativamente, as formas morfológicas preservadas no registro fóssil frequentemente exibem maior complexidade e integração funcional do que suas contrapartes contemporâneas. Este padrão inverte as expectativas evolutivas convencionais e sugere que o registro fóssil representa tipos ancestrais mais complexos, dos quais as formas atuais derivaram através de processos simplificatórios.
Padrões de Estase Morfológica no Registro Fóssil
Táxons em Estase
Proporção de espécies fósseis que não apresentam variação morfológica direcional ao longo de sua distribuição estratigráfica
Aparições Súbitas
Porcentagem de novos táxons que aparecem no registro fóssil sem formas intermediárias documentadas
Transições Graduais
Frequência de sequências evolutivas graduais convincentes no registro paleontológico
Análises quantitativas do registro fóssil revelam que a estase morfológica, não a mudança gradual, constitui o padrão dominante. A vasta maioria dos táxons fósseis mantém morfologia estável ao longo de toda sua distribuição estratigráfica, contradizendo expectativas evolutivas de mudança contínua. Quando mudanças ocorrem, são tipicamente abruptas e não precedidas por formas intermediárias.
Este padrão é observado consistentemente em diferentes grupos taxonômicos, ambientes deposicionais e intervalos temporais, sugerindo que representa uma característica fundamental do registro paleontológico, não um artefato de preservação específica. A predominância de estase morfológica é mais compatível com um modelo de sepultamento catastrófico de tipos estáveis do que com processos evolutivos graduais.
Ausência de Formas Intermediárias
A ausência sistemática de formas intermediárias representa um dos aspectos mais problemáticos do registro fóssil para teorias evolutivas graduais. Apesar de 150 anos de coleta intensiva e milhões de espécimes catalogados, as transições morfológicas graduais permanecem excepcionalmente raras. Esta escassez não pode ser adequadamente explicada por preservação diferencial, pois afeta igualmente grupos com excelente potencial de fossilização.
Grupos com registro fóssil particularmente denso, como foraminíferos, trilobitas e moluscos, deveriam preservar abundantes formas intermediárias se a evolução gradual fosse o mecanismo primário de diversificação. Contudo, mesmo nestes grupos excepcionalmente bem documentados, as transições morfológicas são tipicamente abruptas, sugerindo que o padrão reflete processos biológicos reais, não artefatos tafonômicos.
Complexidade Morfológica nos Fósseis Ancestrais
Uma observação particularmente intrigante no registro paleontológico é que muitos táxons fósseis apresentam maior complexidade morfológica e integração funcional do que seus supostos descendentes evolutivos. Os trilobitas do Cambriano, por exemplo, possuíam sistemas visuais compostos mais sofisticados do que a maioria dos artrópodes contemporâneos. Esta inversão do padrão evolutivo esperado desafia interpretações convencionais.
Análises morfométricas comparativas demonstram que a tendência geral no registro fóssil é de simplificação, não de complexificação. Características morfológicas elaboradas tendem a desaparecer ao longo do tempo, enquanto inovações funcionais genuínas são excepcionalmente raras. Este padrão de degradação morfológica é consistente com um modelo degenerativo de diversificação biológica.
A preservação de sistemas funcionais altamente integrados em táxons fósseis antigos sugere que estes representam tipos morfológicos básicos originais, não formas primitivas em processo de evolução ascendente. A complexidade observada em fósseis ancestrais apoia a interpretação de que o registro paleontológico documenta tipos criados originais, posteriormente diversificados através de processos degenerativos.
Repetições Morfológicas no Registro Fóssil
Morfologias Recorrentes
Padrões morfológicos que se repetem em diferentes intervalos temporais
Arquétipos Básicos
Limitações desenvolvimentais que canalizam a diversificação morfológica
Preservação Seletiva
Sepultamento preferencial de formas morfológicas estáveis
O registro geopaleontológico é caracterizado por repetições morfológicas notáveis – padrões estruturais que reaparecem em diferentes grupos taxonômicos e intervalos temporais. Estas convergências morfológicas são frequentemente interpretadas como evidência de pressões seletivas similares, mas podem refletir limitações desenvolvimentais fundamentais derivadas de arquétipos morfológicos básicos.
A recorrência de padrões morfológicos específicos sugere que a diversificação biológica opera dentro de limitações estruturais definidas, consistente com variação sobre temas morfológicos básicos preexistentes. Esta observação apoia a interpretação de que o registro fóssil documenta amostras de tipos morfológicos fundamentais, dos quais toda diversidade subsequente derivou através de modificações limitadas.
Análise Tafonômica: Sepultamento Catastrófico
Evidências tafonômicas abundantes demonstram que grande parte do registro fóssil resulta de eventos de sepultamento catastrófico, não de processos deposicionais graduais. Fósseis preservados em posições de vida, articulação completa de esqueletos, e preservação de tecidos moles indicam soterramento rápido que impediu decomposição e dispersão normal.
Análises sedimentológicas de camadas fossilíferas revelam características consistentes com deposição turbulenta de alta energia, não com acumulação gradual em ambientes estáveis. A distribuição global de depósitos fossilíferos similares sugere eventos catastróficos de escala continental ou global, incompatíveis com processos uniformitarianistas lentos.
Esta evidência tafonômica apoia a interpretação de que o registro fóssil representa “instantâneos” de biotas ancestrais, preservados durante eventos de sepultamento catastrófico. A preservação excepcional de organismos completos e comunidades ecológicas inteiras é mais compatível com catástrofe súbita do que com fossilização gradual ao longo de eras geológicas.
Evidências de Catástrofe Global Recente
Pico de Mutações Mitocondriais
Evidência genética de gargalo populacional severo na história humana recente
Camadas de Impacto Global
Distribuição worldwide de camadas ricas em irídio e microesferas de impacto
Extinções Megafaunais
Desaparecimento súbito de grandes mamíferos em escala global
Reorganização Geológica
Evidências de atividade tectônica e vulcanismo intensos recentes
Múltiplas linhas de evidência convergem para demonstrar que o planeta Terra experimentou uma catástrofe global de magnitude extraordinária em sua história recente. Dados genéticos indicam gargalos populacionais severos em múltiplas espécies, consistentes com eventos de quase-extinção. O registro arqueológico documenta descontinuidades culturais abruptas e reorganizações populacionais em escala continental.
Evidências geológicas incluem camadas de impacto distribuídas globalmente, anomalias geoquímicas indicativas de atividade vulcanismo extrema, e reorganizações tectônicas recentes. Esta convergência de evidências independentes suporta a ocorrência de uma catástrofe global que teria resultado no sepultamento massivo de biotas preexistentes e na subsequente recolonização por populações sobreviventes reduzidas.
Impactos Catastróficos e Decaimento Radioativo Acelerado
Pesquisas recentes documentam correlações significativas entre eventos de impacto catastróficos e anomalias no decaimento radioativo. Impactos de alta energia podem induzir alterações nas constantes de decaimento nuclear através de efeitos quânticos e modificações no ambiente eletromagnético local. Esta descoberta tem implicações profundas para a interpretação de idades radiométricas convencionais.
Análises de materiais de impacto revelam evidências de aceleração do decaimento radioativo em ordens de magnitude durante eventos catastróficos. Se confirmadas, estas observações invalidariam as cronologias radiométricas baseadas em taxas de decaimento constantes, exigindo uma reavaliação fundamental das escalas temporais geológicas aceitas.
A correlação entre catástrofes e anomalias radiométricas oferece uma explicação alternativa para as aparentes inconsistências cronológicas no registro geopaleontológico. Eventos de sepultamento catastrófico acompanhados por aceleração do decaimento radioativo resultariam em idades aparentes antigas para materiais na realidade recentes, reconciliando evidências geológicas com cronologias históricas.
Crítica às Datações Radiométricas Convencionais
Análises sistemáticas de datações radiométricas revelam inconsistências fundamentais que questionam a confiabilidade das cronologias geológicas convencionais. Inversões estratigráficas, onde rochas supostamente mais antigas sobrepõem rochas mais jovens, são frequentes e não podem ser explicadas adequadamente por processos geológicos normais.
Discordâncias sistemáticas entre diferentes métodos radiométricos aplicados às mesmas amostras indicam problemas fundamentais nas suposições básicas destes métodos. A premissa de taxas de decaimento constantes é particularmente problemática à luz de evidências de variação histórica nas constantes físicas fundamentais.
Geologia da Terra: Evidências de Acidente Global
A análise geológica independente da Terra revela evidências inequívocas de um acidente global de magnitude extraordinária. Formações sedimentares de escala continental, características de deposição turbulenta de alta energia, e distribuições minerais anômalas apontam para eventos catastróficos que transcendem processos geológicos uniformes.
Estruturas geológicas como discordâncias angulares extensas, falhas de escala continental, e reorganizações topográficas dramáticas indicam períodos de atividade tectônica extrema. A preservação de superfícies planálticas extensas sugere que grandes áreas da crosta terrestre foram modificadas simultaneamente, não gradualmente ao longo de eras geológicas.
Depósitos sedimentares globais contendo fósseis marinhos em elevações continentais elevadas fornecem evidência direta de inundação catastrôfica em escala planetária. A distribuição e características destes depósitos são consistentes com um evento diluvial global, não com processos marinhos lentos localizados.
Distribuição Global de Depósitos Diluviais
Fósseis Marinhos Continentais
Descoberta de fauna marinha fossilizada em elevações superiores a 4.000 metros
Camadas Sedimentares Continentais
Formações sedimentares extensas cobrindo milhares de quilômetros quadrados
Depósitos de Carvão Alóctones
Camadas de carvão sem solos desenvolvidos, indicando deposição rápida
A distribuição global de depósitos sedimentares contendo fósseis marinhos em ambientes continentais fornece evidência convincente de inundação catastrôfica planetária. Estes depósitos não podem ser explicados adequadamente por processos marinhos graduais ou transgressões localizadas, pois sua escala transcende qualquer fenômeno oceânico observável contemporaneamente.
A uniformidade litológica e fossilífera destes depósitos sugere deposição sincronizada durante um evento único de magnitude global. A ausência de superfícies de erosão significativas entre camadas indica deposição contínua sob condições energéticas extremas, incompatíveis com ambientes deposicionais estáveis prolongados.
Teoria da Degeneração das Espécies: Fundamentos Conceituais
A Teoria da Degeneração das Espécies (TDE) propõe que toda a diversidade biológica atual derivou de um conjunto limitado de tipos básicos ancestrais através de processos degenerativos de subespeciação. Este modelo inverte a interpretação evolutiva convencional, postulando que a complexidade biológica diminui, não aumenta, ao longo do tempo através de processos genéticos observáveis.
Os mecanismos degenerativos incluem mutações deletérias, recombinação que segrega características ancestrais integradas, deriva genética que fixa alelos deletérios em populações pequenas, e regulação epigenética alterada que reduz a expressão de genes funcionais. Todos estes processos são empiricamente documentados e resultam em redução da informação genética útil.
A TDE oferece explicações parcimoniosas para padrões observados tanto no registro fóssil quanto em populações contemporâneas, integrando dados paleontológicos, genéticos e observacionais em um framework teórico coerente que evita as contradições inerentes aos modelos evolutivos ascendentes convencionais.
Mecanismos Degenerativos de Diversificação
Mutações Deletérias
Alterações genéticas que reduzem ou eliminam funções gênicas preexistentes
Recombinação Segregativa
Separação de alelos funcionais anteriormente integrados em sistemas genéticos coordenados
Deriva Genética
Fixação aleatória de variantes deletérias em populações de tamanho efetivo reduzido
Desregulação Epigenética
Alterações na expressão gênica que reduzem a produção de proteínas funcionais
Os mecanismos degenerativos operam através de processos genéticos bem caracterizados que resultam consistentemente na perda de informação genética funcional. Mutações deletérias são estatisticamente mais prováveis que mutações benéficas, criando uma pressão degenerativa constante sobre genomas complexos. A recombinação, embora essencial para reprodução sexual, fragmenta sistemas genéticos integrados acumulados em tipos ancestrais.
Deriva genética em populações pequenas fixa mutações deletérias independentemente de seus efeitos fenotípicos, acelerando a degradação genômica. Modificações epigenéticas podem silenciar genes funcionais sem alterar sequências de DNA, produzindo variação fenotípica através de perda funcional reversível. Estes mecanismos fornecem bases genéticas robustas para o modelo degenerativo de diversificação.
Evidências Moleculares da Degeneração Genômica
Análises genômicas comparativas fornecem evidência molecular direta para processos degenerativos em andamento. Comparações entre genomas de espécies relacionadas revelam padrões consistentes de perda gênica, pseudogenização, e redução da complexidade regulatória. Estes padrões são observados em múltiplos táxons e ambientes, sugerindo um processo universal de degradação genômica.
Estudos de genomas organelares demonstram taxas elevadas de mutação deletéria e perda funcional em linhagens derivadas comparadas a ancestrais inferidos. A acumulação de mutações deletérias excede significativamente a fixação de mutações benéficas, resultando em degradação líquida da informação genética ao longo do tempo evolutivo.
Análises filogenômicas indicam que a complexidade genômica tende a diminuir nas pontas das árvores filogenéticas, onde espécies derivadas possuem genomas menores e menos complexos que seus ancestrais reconstruídos. Esta observação contradiz expectativas evolutivas de complexificação progressiva e apoia fortemente o modelo degenerativo proposto.
Ausência de Complexidade Molecular Progressiva Crescente (CMPCCN)
Inovações Funcionais
Proporção de mutações que genuinamente aumentam complexidade funcional
Mutações Neutras
Alterações sem efeito funcional detectável nos sistemas biológicos
Mutações Deletérias
Modificações que reduzem ou eliminam funções biológicas preexistentes
A análise sistemática de mutações moleculares revela a ausência virtual de Complexidade Molecular Progressiva Crescente de Novo (CMPCCN) – o surgimento genuíno de nova informação genética funcional. A vasta maioria das mutações observadas são deletérias ou neutras, com uma fração mínima conferindo vantagem funcional genuína.
Mesmo mutações classificadas como “benéficas” frequentemente representam otimizações de funções preexistentes ou ativação de capacidades latentes, não criação de nova informação genética. A raridade extrema de inovações funcionais genuínas contrasta dramaticamente com as expectativas de modelos evolutivos que requerem geração contínua de nova informação genética para explicar a diversidade biológica observada.
Esta assimetria fundamental entre perda e ganho de informação genética fornece evidência molecular robusta para o modelo degenerativo. A predominância de processos que degradam informação genética sobre aqueles que a criam é incompatível com evolução ascendente sustentada e apoia fortemente processos degenerativos como mecanismo primário de diversificação biológica.
Onipresença de Complexidade Degradante Molecular Atual (CDMA)
Em contraste com a raridade de CMPCCN, observamos onipresença de Complexidade Degradante Molecular Atual (CDMA) – processos que consistentemente reduzem a informação genética funcional. CDMA é documentada em todas as linhagens estudadas e representa o padrão molecular dominante em sistemas biológicos contemporâneos.
Mecanismos de CDMA incluem acúmulo de mutações deletérias em genes funcionais, perda de elementos regulatórios que coordenam expressão gênica, e fragmentação de operons e clusters genéticos funcionalmente integrados. Estes processos operam continuamente e são detectáveis mesmo em populações grandes com seleção natural eficiente.
A universalidade de CDMA indica que todos os organismos contemporâneos estão sujeitos a degradação genômica contínua. Esta observação é inconsistente com modelos evolutivos que postulam melhoramento genético líquido ao longo do tempo, mas é perfeitamente compatível com diversificação degenerativa a partir de tipos ancestrais mais complexos preservados no registro fóssil.
Comparação Quantitativa: Tempo Real vs. Tempo Geológico
A análise quantitativa comparativa revela uma discrepância fundamental entre as taxas de variação morfológica observadas em diferentes escalas temporais. Processos de subespeciação em tempo real e arqueológico operam ordens de magnitude mais rapidamente do que previsto por modelos evolutivos graduais, enquanto o registro geopaleontológico apresenta taxas de mudança drasticamente menores que as expectativas teóricas.
Esta inversão de expectativas temporais não pode ser explicada adequadamente por modelos evolutivos convencionais, que predizem correlação positiva entre tempo disponível e magnitude da mudança morfológica. A observação de variação rápida em escalas curtas e estase em escalas longas sugere processos fundamentalmente diferentes operando nestas escalas temporais distintas.
Implicações para Cronologias Evolutivas
Subespeciação Rápida
Variação morfológica documentada em décadas a milênios
Lacuna Temporal
Ausência de processos intermediários entre escalas observáveis e geológicas
Estase Geopaleontológica
Ausência de variação direcional no registro fóssil
A discrepância temporal fundamental entre subespeciação observável e estase geopaleontológica tem implicações profundas para cronologias evolutivas convencionais. Se processos contemporâneos operassem uniformemente ao longo de eras geológicas, deveríamos observar variação morfológica extrema no registro fóssil, não a estase documentada.
Alternativamente, se as escalas temporais geológicas convencionais fossem corretas, os processos de subespeciação contemporâneos deveriam operar muito mais lentamente para serem compatíveis com os padrões paleontológicos observados. A incompatibilidade entre estas escalas temporais sugere problemas fundamentais com um ou ambos os conjuntos de observações.
O modelo degenerativo resolve esta contradição propondo que a subespeciação contemporânea representa diversificação pós-catastrófica recente a partir de tipos básicos preservados no registro fóssil, eliminando a necessidade de extrapolação de processos através de escalas temporais incompatíveis.
Tipos Básicos Originais: Definição e Características
Tipos básicos originais representam organismos criados com alta complexidade genômica e morfológica, caracterizados por sistemas funcionais altamente integrados e coordenados. Estes organismos possuíam genomas complexos contendo informação genética suficiente para gerar toda a diversidade subsequente observada em seus grupos descendentes através de processos de variação controlada.
Características distintivas incluem alta redundância funcional, sistemas regulatórios complexos, e capacidade latente para expressar fenótipos alternativos sob diferentes condições ambientais. Esta plasticidade fenotípica permitiu colonização de ambientes diversos após eventos catastróficos, mas às custas de perda progressiva de informação genética através de especialização.
O conceito de tipos básicos originais é operacionalmente definido através da identificação de morfotipos no registro fóssil que exibem maior complexidade e integração funcional que suas contrapartes contemporâneas, sugerindo que representam formas ancestrais das quais toda diversidade atual derivou através de processos simplificatórios.
Evidências de Integração Funcional em Fósseis Ancestrais
Sistemas Visuais Complexos
Trilobitas com olhos compostos mais sofisticados que artrópodes modernos, demonstrando maior integração sensorial
Arquiteturas Esqueletárias
Amonites com padrões de sutura complexos ausentes em cefalópodes contemporâneos
Sistemas Locomotores
Dinossauros com integração biomecânica superior a qualquer vertebrado terrestre atual
Análises morfológicas comparativas de táxons fósseis revelam níveis de integração funcional frequentemente superiores aos observados em organismos contemporâneos. Esta observação contraintuitiva desafia interpretações evolutivas convencionais que predizem aumento de complexidade ao longo do tempo geológico.
Sistemas funcionais integrados em fósseis ancestrais demonstram coordenação entre múltiplos sistemas orgânicos que é raramente igualada em formas contemporâneas. Esta maior integração sugere que organismos ancestrais possuíam sistemas de desenvolvimento mais coordenados e genomas mais integrados que seus supostos descendentes evolutivos.
Sepultamento e Preservação de Populações Ancestrais
O registro geopaleontológico documenta extensively eventos de sepultamento em massa que preservaram populações inteiras de organismos ancestrais. Estes eventos são caracterizados por preservação excepcional de organismos completos, manutenção de articulação esquelética, e conservação de tecidos moles – características incompatíveis com processos tafonômicos graduais normais.
Análises de assembleias fósseis revelam preservação de comunidades ecológicas completas, incluindo organismos de diferentes níveis tróficos e habitats ecológicos. Esta preservação holística de ecosistemas inteiros indica sepultamento catastrófico simultâneo de biotas preexistentes, não acumulação gradual de restos organísticos ao longo de períodos extensos.
A distribuição estratigráfica de eventos de preservação excepcional correlaciona-se com evidências sedimentológicas de deposição catastrófica, sugerindo que o registro fóssil representa principalmente “instantâneos” de biotas ancestrais preservados durante eventos globais de sepultamento em massa. Esta interpretação explica tanto a qualidade excepcional da preservação quanto a estase morfológica característica do registro paleontológico.
Lagerstätten e Preservação Excepcional
Folhelho de Burgess
Preservação de fauna de tecidos moles demonstrando diversidade morfológica extrema e sistemas funcionais complexos
Calcário de Solnhofen
Conservação excepcional de vertebrados completos com preservação de estruturas delicadas como penas
Explosão Cambriana
Aparição súbita de planos corporais complexos sem precursores evolutivos documentados
Lagerstätten (depósitos de preservação excepcional) fornecem evidência direta de sepultamento catastrófico de populações ancestrais. A qualidade extraordinária da preservação nestas localidades requer condições deposicionais excepcionais que impedem decomposição e desarticulação normais. Tais condições são incompatíveis com ambientes deposicionais estáveis prolongados postulados por modelos uniformitarianistas.
A diversidade morfológica e complexidade funcional documentadas em Lagerstätten frequentemente excedem aquelas observadas em ecosistemas contemporâneos comparáveis. Esta observação sugere que as faunas preservadas representam tipos básicos originais com maior complexidade e diversidade morfológica que suas contrapartes modernas degradadas.
Padrões de Radiação Pós-Catastrófica
Evento Catastrófico
Sepultamento massivo de biotas ancestrais e redução populacional drástica
Populações Refugiais
Sobrevivência de pequenos grupos em ambientes protegidos
Recolonização
Expansão geográfica e ocupação de nichos ecológicos vagos
Diferenciação Degenerativa
Subespeciação através de deriva genética e perda de informação
O modelo degenerativo prediz padrões específicos de radiação pós-catastrófica que são amplamente observados no registro biológico. Populações pequenas sobreviventes experimentam gargalos genéticos severos que promovem deriva genética e fixação de mutações deletérias. A subsequente expansão populacional e geográfica resulta em diferenciação através de processos degenerativos, não inovação evolutiva.
Evidências genéticas de gargalos populacionais recentes em múltiplas espécies são consistentes com eventos catastróficos globais que reduziram drasticamente o tamanho populacional efetivo. A diversidade genética reduzida observada em muitas espécies contemporâneas reflete estes gargalos históricos e os processos degenerativos subsequentes.
Biogeografia e Padrões de Dispersão Pós-Diluvial
Padrões biogeográficos globais exibem características consistentes com dispersão pós-catastrófica a partir de áreas refugiais limitadas. A distribuição de espécies relacionadas em continentes separados pode ser explicada através de migração recente facilitada por pontes terrestres expostas durante períodos de nível marinho reduzido pós-catástrofe.
Análises filogenéticas de espécies com distribuição disjunta revelam divergência recente incompatível com separação continental antiga. Estes padrões sugerem colonização rápida de continentes a partir de populações fundadoras pequenas, seguida por diferenciação degenerativa in situ através de processos de subespeciação local.
A concentração de diversidade morfológica em regiões específicas (hotspots de biodiversidade) é consistente com radiação a partir de centros de dispersão pós-catastrófica. Estas regiões funcionaram como “arcas” biogeográficas onde populações ancestrais sobreviveram ao evento catastrófico global e subsequentemente irradiaram para recolonizar áreas devastadas.
Centros de Diversificação e “Arcas” Biogeográficas
Regiões Montanhosas
Elevações ofereceram refúgio durante inundações catastróficas
Florestas Tropicais
Ambientes estáveis que preservaram diversidade ancestral
Sistemas Insulares
Ilhas funcionaram como refugia isolados para populações ancestrais
Oásis e Vales
Microambientes protegidos em regiões áridas
Sistemas Cavernícolas
Ambientes subterrâneos ofereceram proteção total
Regiões de alta biodiversidade contemporânea correspondem geograficamente a áreas que teriam oferecido refúgio durante eventos catastróficos globais. Estas “arcas” biogeográficas preservaram núcleos de diversidade ancestral que subsequentemente irradiaram através de processos degenerativos de subespeciação para recolonizar ambientes devastados pela catástrofe.
A correlação entre topografia refugial e diversidade contemporânea não é explicada adequadamente por modelos evolutivos convencionais, que predizem diversificação em resposta a oportunidades ecológicas, não conservação de diversidade preexistente. O padrão observado é mais consistente com preservação e subsequente irradiação de tipos ancestrais diversos.
Degeneração Genética em Populações Isoladas
Populações isoladas fornecem experimentos naturais de degeneração genética que validam empiricamente o modelo degenerativo. Espécies endêmicas de ilhas consistentemente exibem características degenerativas: redução do tamanho corporal, perda de capacidade de voo, simplificação de sistemas sensoriais, e redução da diversidade genética comparada a populações ancestrais continentais.
Análises genômicas de espécies insulares revelam acúmulo de mutações deletérias, perda de diversidade alélica, e redução da eficiência da seleção natural devido a tamanhos populacionais pequenos. Estes processos operam em escalas temporais observáveis, fornecendo analogias diretas para processos degenerativos em escalas temporais maiores.
A universalidade de características degenerativas em populações isoladas demonstra que a degeneração genética é um processo inevitável em populações de tamanho reduzido. Esta observação apoia a interpretação de que a diversidade biológica contemporânea deriva de populações ancestrais que experimentaram gargalos severos durante eventos catastróficos globais.
Síndromes de Insularidade como Modelo Degenerativo
Nanismo Insular
Redução sistemática do tamanho corporal em espécies insulares através de perda de fatores de crescimento
Perda de Capacidade de Voo
Atrofia de sistemas de voo em aves e insetos isolados, representando perda funcional
Simplificação Sensorial
Redução de sistemas visuais e auditivos em ambientes com predação reduzida
Redução Comportamental
Perda de comportamentos complexos ancestrais desnecessários em ambientes simplificados
Síndromes de insularidade representam manifestações aceleradas de processos degenerativos que operam universalmente, mas são mais evidentes em populações isoladas. A previsibilidade destes processos permite testar empiricamente as predições do modelo degenerativo em escalas temporais observáveis.
Todas as características da síndrome de insularidade representam perdas ou simplificações de características ancestrais, não inovações evolutivas. Esta observação consistente em múltiplos táxons e ambientes insulares fornece evidência robusta para processos degenerativos como mecanismo primário de diversificação biológica.
Domesticação e Aceleração da Degeneração
A domesticação animal representa o experimento mais extenso de subespeciação dirigida disponível para análise científica. Em escalas temporais extremamente curtas (menos de 15.000 anos), a seleção artificial produziu diversidade morfológica que rivaliza com a observada entre espécies selvagens relacionadas. Esta radiação extrema demonstra o potencial latente dos genomas ancestrais para gerar diversidade através de processos degenerativos acelerados.
Análises genômicas de espécies domésticas revelam que praticamente toda a diversidade morfológica resulta de mutações deletérias, perda de função regulatória, e segregação de alelos ancestrais. A seleção artificial fixou mutações que seriam eliminadas pela seleção natural, acelerando processos degenerativos que normalmente operariam mais lentamente.
A morfologia extrema de muitas raças domésticas (cães braquicefálicos, gatos sem pelos, etc.) ilustra as consequências de processos degenerativos descontrolados. Estas formas representam o endpoint lógico da degeneração genética, demonstrando que processos degenerativos podem produzir diversidade morfológica extrema, mas às custas da integridade funcional.
Comparação Genômica: Espécies Ancestrais vs. Derivadas
Análises genômicas comparativas sistemáticas demonstram que espécies derivadas consistentemente possuem genomas menores, menos genes funcionais, elementos regulatórios reduzidos, e diversidade alélica diminuída comparadas a espécies ancestrais inferidas. Esta redução sistemática da complexidade genômica é incompatível com modelos evolutivos ascendentes.
A correlação negativa entre posição filogenética derivada e complexidade genômica indica que processos degenerativos operam consistentemente ao longo da diversificação biológica. Espécies nas pontas de árvores filogenéticas (mais derivadas) possuem genomas sistematicamente degradados comparados a seus ancestrais reconstruídos.
Pseudogenização e Perda Funcional Progressiva
Gene Funcional Original
Sequência codificante intacta com função biológica específica
Mutações Deletérias
Acúmulo de substituições que reduzem eficiência funcional
Pseudogene Funcional
Função residual mantida mas significativamente reduzida
Pseudogene Não-Funcional
Perda completa de função biológica detectável
Deleção Genômica
Eliminação física da sequência do genoma
O processo de pseudogenização representa um modelo molecular claro de degeneração genética progressiva. Genes funcionais acumulam mutações deletérias que gradualmente reduzem sua eficiência até perda completa de função. Este processo é irreversível e universalmente observado em genomas sequenciados, fornecendo evidência molecular direta de degeneração genômica em andamento.
Análises de genomas relacionados revelam que espécies derivadas possuem mais pseudogenes que espécies ancestrais, indicando aceleração da pseudogenização em linhagens mais recentes. Esta observação é consistente com degradação genômica progressiva ao longo do tempo evolutivo, contrária às expectativas de modelos evolutivos ascendentes.
Refutação de Objeções Evolutivas Convencionais
Críticos do modelo degenerativo frequentemente argumentam que processos evolutivos convencionais podem explicar adequadamente os padrões observados sem invocar degeneração sistemática. Entretanto, análises quantitativas rigorosas demonstram que objeções evolutivas padrão são inadequadas para explicar a magnitude e direção das mudanças morfológicas observadas.
A objeção de “tempo insuficiente” para processos evolutivos convencionais é particularmente problemática à luz de evidências de subespeciação rápida. Se organismos podem diversificar morfologicamente em escalas temporais históricas, por que o registro geopaleontológico não documenta variação comparável? A discrepância temporal sugere problemas fundamentais com interpretações evolutivas convencionais.
Objeções baseadas em “incompletude do registro fóssil” também são inadequadas, pois o padrão de estase e aparições súbitas persiste mesmo em sequências sedimentares com preservação excepcional. A qualidade da preservação em muitos depósitos fossilíferos é suficiente para documentar variação morfológica se esta ocorresse, mas tal variação está consistentemente ausente.
Inadequação da Síntese Evolutiva Moderna
Problema das Escalas Temporais
Incompatibilidade entre taxas de mudança observadas e previstas pela teoria evolutiva
Ausência de Inovação Molecular
Raridade extrema de mutações que genuinamente aumentam informação genética funcional
Estase Paleontológica
Persistência de formas morfológicas sem mudança direcional por períodos geologicamente extensos
Complexidade Fóssil
Maior integração funcional em táxons fósseis comparados a formas contemporâneas
A síntese evolutiva moderna enfrenta contradições fundamentais que não podem ser resolvidas através de modificações incrementais da teoria. Os problemas identificados representam falhas conceituais básicas que exigem reconsideração fundamental dos paradigmas interpretativos em biologia evolutiva.
Tentativas de reconciliar evidências contraditórias através de hipóteses ad hoc (equilíbrios pontuados, constrações desenvolvimentais, etc.) resultam em modelos cada vez mais complexos e menos parcimoniosos. O modelo degenerativo oferece explicações mais diretas e testáveis para os padrões observados.
Evidências Convergentes para o Modelo Degenerativo
A força do modelo degenerativo reside na convergência de evidências independentes de múltiplas disciplinas científicas. Dados paleontológicos, genéticos, biogeográficos, e observacionais contemporâneos convergem para apoiar processos degenerativos como mecanismo primário de diversificação biológica. Esta convergência multidisciplinar não é observada para modelos evolutivos convencionais.
Evidências paleontológicas demonstram estase morfológica e complexidade superior em formas ancestrais. Dados genéticos revelam predominância de processos degenerativos sobre inovação funcional. Padrões biogeográficos são consistentes com radiação pós-catastrófica. Observações contemporâneas documentam subespeciação através de perda de informação genética.
A integração coerente destas evidências diversas em um framework explicativo único demonstra a superioridade do modelo degenerativo sobre interpretações evolutivas fragmentárias que requerem hipóteses ad hoc para explicar anomalias. O poder explicativo unificado do modelo degenerativo constitui evidência forte para sua validade científica.
Predições Testáveis do Modelo Degenerativo
Redução Genômica Progressiva
Espécies derivadas devem possuir genomas menores e menos complexos que ancestrais
Aceleração da Pseudogenização
Taxa de perda gênica deve exceder taxa de ganho funcional em todas as linhagens
Correlação Morfológica-Temporal
Complexidade morfológica deve correlacionar negativamente com posição filogenética derivada
Gargalos Genéticos Universais
Todas as espécies devem exibir evidências de redução populacional histórica recente
Ausência de CMPCCN
Inovações funcionais genuínas devem ser estatisticamente raras em todas as escalas temporais
O modelo degenerativo gera predições específicas e testáveis que podem ser avaliadas através de pesquisas futuras. Estas predições contrastam claramente com expectativas de modelos evolutivos convencionais, permitindo discriminação empírica entre paradigmas alternativos.
A testabilidade das predições degenerativas através de metodologias genômicas, paleontológicas, e experimentais contemporâneas garante que o modelo pode ser rigorosamente avaliado. Confirmação ou refutação destas predições fornecerá evidência definitiva para a validade do paradigma degenerativo.
Metodologias para Teste Empírico
Genômica Comparativa
Sequenciamento de genomas completos de espécies ancestrais e derivadas para análise de complexidade
Análise Filogenômica
Reconstrução de características genômicas ancestrais e direções de mudança evolutiva
Experimentos de Mutação
Caracterização quantitativa de efeitos funcionais de mutações aleatórias
Análise Morfométrica
Quantificação de complexidade morfológica em séries temporais paleontológicas
Metodologias avançadas de genômica comparativa, análise filogenômica, e morfometria quantitativa fornecem ferramentas poderosas para testar empiricamente as predições do modelo degenerativo. Estas metodologias permitem análises quantitativas rigorosas que transcendem argumentos qualitativos tradicionais em biologia evolutiva.
A disponibilidade crescente de genomas completos e técnicas analíticas avançadas cria oportunidades sem precedentes para avaliação empírica definitiva do modelo degenerativo. Resultados preliminares usando estas metodologias já demonstram suporte substancial para predições degenerativas.
Implicações para Políticas de Conservação
O modelo degenerativo tem implicações práticas importantes para estratégias de conservação biológica. Se a diversidade biológica resulta de degeneração progressiva de tipos ancestrais mais complexos, então a conservação deve priorizar a preservação de informação genética ancestral remanescente, não a facilitação de “evolução” futura.
Populações pequenas são particularmente vulneráveis à aceleração de processos degenerativos através de deriva genética. Estratégias de conservação devem focar na manutenção de tamanhos populacionais efetivos grandes para minimizar a fixação de mutações deletérias e preservar diversidade genética ancestral.
Programas de breeding em cativeiro devem evitar seleção que promova características extremas, pois estas frequentemente resultam de mutações deletérias que reduzem fitness geral. A preservação de fenótipos “selvagens” mantém a integridade genética ancestral e previne degradação adicional.
Estratégias de Conservação Genética
Preservação de Genomas Intactos
Priorização de populações com menor degradação genética e maior diversidade alélica ancestral
Prevenção de Gargalos
Manutenção de tamanhos populacionais efetivos para minimizar deriva genética deletéria
Conectividade Genética
Facilitação de fluxo gênico para reverter efeitos de deriva genética localizada
Estratégias de conservação informadas pelo modelo degenerativo enfatizam a preservação de integridade genética ancestral como objetivo primário. Esta abordagem contrasta com estratégias evolutivas convencionais que podem inadvertidamente acelerar processos degenerativos através de fragmentação populacional e seleção artificial.
Direções para Pesquisas Futuras
Genômica de População
Análise detalhada de processos degenerativos em populações naturais contemporâneas
Paleogenômica
Sequenciamento de DNA antigo para reconstrução direta de genomas ancestrais
Evolução Experimental
Estudos controlados de processos degenerativos em condições laboratoriais
Avenidas promissoras para pesquisa futura incluem aplicação de técnicas genômicas avançadas para caracterização detalhada de processos degenerativos, paleogenômica para reconstrução direta de genomas ancestrais, e evolução experimental para teste controlado de predições degenerativas.
O desenvolvimento contínuo de metodologias analíticas oferece oportunidades sem precedentes para avaliação rigorosa do modelo degenerativo. Resultados destas pesquisas determinarão definitivamente a validade do paradigma degenerativo versus modelos evolutivos convencionais.
Tecnologias Emergentes e Oportunidades de Pesquisa
Sequenciamento de Terceira Geração
Tecnologias de leitura longa para montagem completa de genomas ancestrais complexos
Genômica de Célula Única
Análise de variação genética em populações com resolução celular individual
DNA Antigo
Recuperação de sequências genéticas de espécimes fósseis preservados
Tecnologias emergentes em genômica fornecem ferramentas cada vez mais poderosas para investigação direta de processos degenerativos. Sequenciamento de DNA antigo, em particular, oferece acesso direto a genomas ancestrais para comparação com formas derivadas contemporâneas.
Síntese das Evidências e Conclusões Principais
A análise comprehensiva apresentada neste estudo demonstra convergência robusta de evidências multidisciplinares em favor do modelo degenerativo de diversificação biológica. Dados paleontológicos, genéticos, biogeográficos, e observacionais contemporâneos apoiam consistentemente processos degenerativos como mecanismo primário de subespeciação, contrastando com predições de modelos evolutivos convencionais.
A discrepância fundamental entre subespeciação rápida observável em tempo real e estase no registro geopaleontológico não pode ser adequadamente explicada por teorias evolutivas graduais. O modelo degenerativo resolve esta contradição propondo que o registro fóssil representa tipos básicos ancestrais sepultados catastroficamente, dos quais toda diversidade atual derivou através de processos simplificatórios pós-catástrofe.
Evidências convergentes de múltiplas disciplinas científicas fornecem suporte empirical robusto para o paradigma degenerativo. A superioridade explicativa deste modelo sobre interpretações evolutivas convencionais justifica sua consideração séria como framework alternativo para compreensão da origem e diversificação da vida na Terra.
Validação Empírica do Modelo Degenerativo
Suporte Paleontológico
Proporção de táxons fósseis que exibem estase morfológica consistente com sepultamento catastrófico
Evidência Genômica
Porcentagem de linhagens que demonstram redução de complexidade genômica ao longo do tempo
Padrões Biogeográficos
Consistência de distribuições de espécies com radiação pós-catastrófica
Análises quantitativas sistemáticas demonstram suporte empírico extraordinariamente forte para predições do modelo degenerativo. A convergência de evidências independentes de múltiplas disciplinas científicas raramente observada em biologia evolutiva fornece validação robusta do paradigma degenerativo.
O contraste entre o suporte empírico consistente para o modelo degenerativo e as contradições internas de teorias evolutivas convencionais sugere que uma mudança de paradigma fundamental pode ser necessária para progresso científico em biologia evolutiva. O modelo degenerativo oferece esta alternativa teoricamente coerente e empiricamente suportada.
Implicações Filosóficas e Metodológicas
A validação do modelo degenerativo tem implicações profundas para a filosofia da ciência em biologia evolutiva. A predominância de processos degenerativos sobre inovação evolutiva sugere que a complexidade biológica é primordial, não emergente, invertendo suposições fundamentais sobre a natureza da vida e seu desenvolvimento ao longo do tempo.
Metodologicamente, a aceitação do modelo degenerativo exigiria reorientação fundamental das prioridades de pesquisa em biologia evolutiva. Em vez de buscar mecanismos de inovação evolutiva, esforços científicos deveriam focar na compreensão de processos degenerativos e estratégias para sua mitigação ou reversão.
A mudança paradigmática proposta não representa meramente uma modificação teórica, mas uma revolução conceitual comparável às grandes transições na história da ciência. Como todas as revoluções científicas, requer reavaliação fundamental de evidências existentes e reformulação de questões de pesquisa básicas.
Recomendações para a Comunidade Científica
Reavaliação de Evidências
Análise sistemática de dados existentes sob a perspectiva do modelo degenerativo
Testes Empíricos Diretos
Implementação de experimentos específicos para discriminar entre modelos alternativos
Reformulação Teórica
Desenvolvimento de frameworks conceituais baseados em processos degenerativos
Diálogo Interdisciplinar
Integração de perspectivas de paleontologia, genética, e biologia de sistemas
Recomendamos que a comunidade científica considere seriamente as evidências apresentadas e implemente programas de pesquisa para teste rigoroso do modelo degenerativo. A magnitude das implicações potenciais justifica investigação intensiva, independentemente de predisposições teóricas prévias.
O progresso científico requer abertura para mudanças paradigmáticas quando evidências empíricas as suportam. A convergência de evidências multidisciplinares para o modelo degenerativo representa precisamente tal situação, exigindo resposta apropriada da comunidade científica internacional.
Conclusões Finais: Um Novo Paradigma para a Biologia Evolutiva
Este estudo demonstra que a subespeciação ocorre exclusivamente em tempo real e arqueológico através de processos degenerativos, estando totalmente ausente do registro geopaleontológico que documenta tipos básicos ancestrais sepultados catastroficamente. As evidências convergentes de múltiplas disciplinas científicas suportam inequivocamente a Teoria da Degeneração das Espécies como framework explicativo superior aos modelos evolutivos convencionais.
O registro fóssil representa amostras de sepultamento de populações ancestrais – tipos básicos originais dotados de alta complexidade genômica e morfológica, dos quais toda a biodiversidade atual derivou através de processos simplificatórios de subespeciação. Esta interpretação resolve as contradições fundamentais entre escalas temporais observáveis e geopaleontológicas que há muito tempo desafiam a biologia evolutiva.
A validação empírica do modelo degenerativo representa uma mudança paradigmática fundamental que requer reavaliação completa de conceitos básicos em biologia evolutiva. Esta revolução científica oferece oportunidades sem precedentes para avanços teóricos e aplicações práticas em conservação, medicina, e biotecnologia. O futuro da biologia evolutiva reside na compreensão e mitigação de processos degenerativos, não na busca ilusória de inovação evolutiva ascendente.
Deixe um comentário