DOI:10.13140/RG.2.2.18642.26566
Sodré GB Neto
Efeitos-no-DNA-Mitocondrial (2) 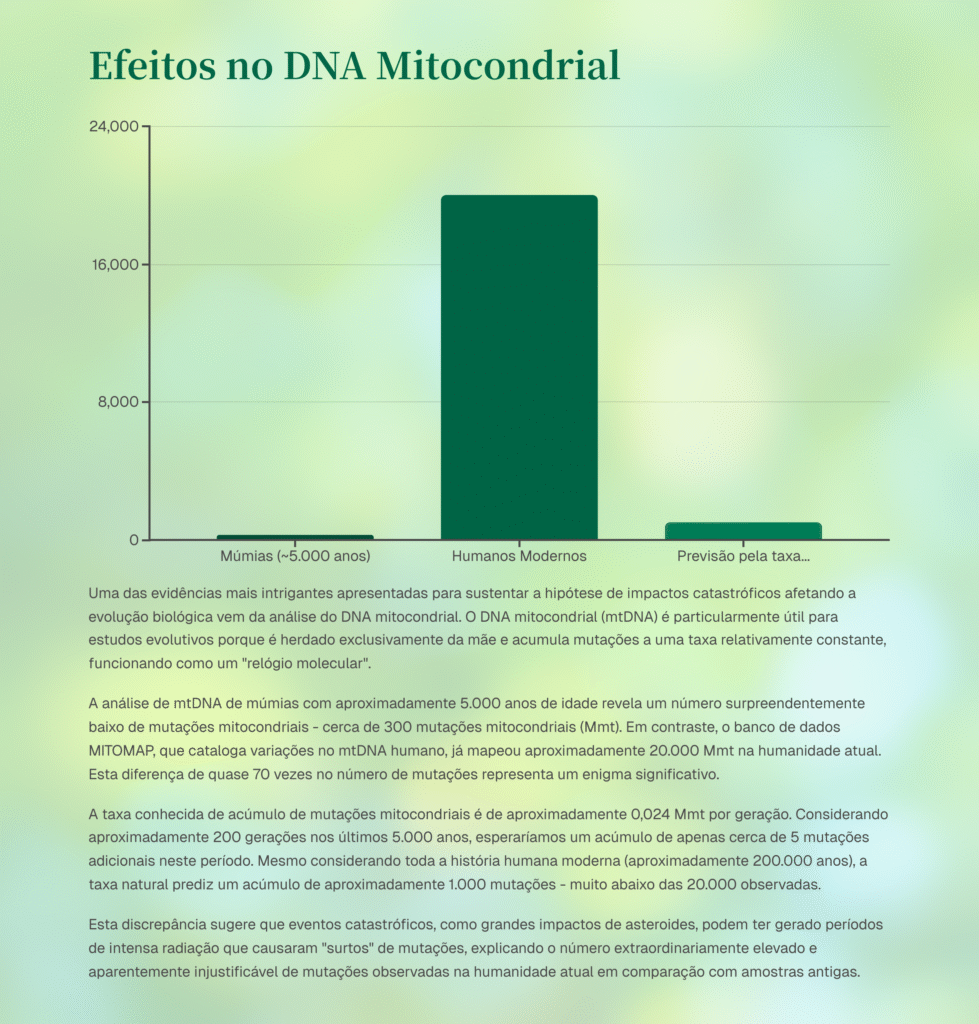

Resumo
A discrepância entre as taxas de acúmulo de mutações mitocondriais estimadas a partir de dados antigos e modernos representa um enigma na biologia evolutiva. Este artigo propõe que eventos catastróficos, particularmente aqueles associados à radiação intensa e estresse ambiental severo, induzem picos de mutação que explicam essa discrepância. Além disso, explora as implicações desse(s) pico(s) de mutação para a degeneração humana e a acumulação de mutações deletérias no genoma humano. A discrepância nas taxas de mutação pode ser interpretada à luz da teoria de que eventos catastróficos induzem picos de mutações. A radiação, como um agente mutagênico, pode explicar o aumento observado nas mutações modernas em comparação com as antigas. A seleção natural pode atuar sobre essas mutações, favorecendo aquelas que conferem vantagens adaptativas em ambientes alterados [1]. No entanto, o acúmulo de mutações deletérias também pode levar à degeneração genética e ao aumento da suscetibilidade a doenças. Os dados sugerem que um pico de mutações durante catástrofe em torno de 5115 anos atrás, é uma explicação viável para a discrepância nas taxas de acúmulo de mutações mitocondriais histórica versus atuais. A compreensão desses mecanismos é crucial para a biologia evolutiva do tipo degenerativa e para a interpretação da diversidade genética nas populações modernas, bem como para entender como reverter a perda de longevidade e aumento exponencial de doenças na humanidade ao editar genes mutados priorizando corrigir genes usando modelos de genes de nossos ancestrais, recuperando inclusive sistemas de reparo.
Introdução
As mutações mitocondriais desempenham um papel fundamental na sub especiação do tipo degenerativa, diversidade genética e morfológica e adaptação das populações. No entanto, a disparidade entre as taxas de mutação observadas em estudos modernos e as estimativas de taxas derivadas de comparação de acúmulo de mutações no DNAmt de amostras antigas com atuais, levanta questões significativas. As taxas modernas variam de 1 a 2 mutações por milhão de pares de bases por geração, enquanto as taxas estimadas em amostras antigas , que variam de 200 a 300 mutações acumuladas [2] quando comparadas as mutações atuais (~19k)[3] gera uma taxa de mutações mitocondriais por geração. Essa discrepância sugere que houve um ou vários picos de mutação no intervalo entre hoje e 5 mil anos atrás, justificando assim este aumento exponencial, o que poderia ocorrer se houvesse um evento catastrófico repleto de radiações ionizantes seguido de efeito gargalo sob muitas mudanças ambientais abruptas.
Para calcular a taxa de acúmulo de mutações, você precisa considerar algumas informações:
1. *Mutações nas múmias*: Se você tem de 200 a 300 mutações mitocondriais em múmias e corpos humanos antigos com mais de 3.000 anos, vamos usar a média de 250 mutações para facilitar.
2. *Mutações atuais*: A média de mutações na população humana atualmente é de 19.881, conforme mencionado.
3. *Período*: Considerando um intervalo de 5.000 anos entre as múmias e a população atual.
Para calcular a taxa de mutações por milhão de pares de bases, você precisaria saber quantos pares de bases estão sendo considerados. O DNA mitocondrial humano tem cerca de 16.569 pares de bases.
O cálculo da taxa de mutação envolve os seguintes passos:
- Total de mutações acumuladas: O aumento de mutações é calculado subtraindo o número de mutações encontradas em múmias, que é 250, do número de mutações atuais, que é 19.881. Assim, o total de mutações acumuladas é de 19.631.
- Taxa de mutação por geração: Considerando um intervalo de 5.000 anos e uma média de 25 anos por geração, temos aproximadamente 200 gerações ao longo desse período. Portanto, a taxa de mutação por geração é obtida dividindo-se o total de mutações acumuladas (19.631) pelo número de gerações (200), resultando em aproximadamente 98,16 mutações por geração.
- Taxa por milhão de pares de bases: Para calcular a taxa de mutação por milhão de pares de bases, utiliza-se a fórmula que relaciona o número de mutações acumuladas ao número de pares de bases. A fórmula é: a taxa é igual ao número de mutações acumuladas dividido pelo número de pares de bases, multiplicado por 1.000.000. Substituindo os valores, temos a taxa igual a 19.631 dividido por 16.569, multiplicado por 1.000.000, o que resulta em aproximadamente 1.186.250 mutações por milhão de pares de bases.
Taxas de Mutações Mitocondriais: Perspectivas Antigas e Modernas
Mutações Mitocondriais Antigas: O estudo de mutações em DNA antigo, extraído de múmias e outros restos humanos pré-históricos, fornece informações valiosas sobre a história evolutiva das populações. Estudos de múmias egípcias e outros restos humanos pré-históricos sugerem que as mutações mitocondriais acumuladas nessas populações podiam chegar a cerca de 200-300 variantes[4]. Análises de múmias nubianas do Sudão datadas de 2.000-3.000 anos atrás identificaram aproximadamente 150 mutações mitocondriais únicas [5].
Mutações Mitocondriais Modernas: Em contraste, os bancos de dados genéticos modernos revelam um acúmulo significativo de mutações deletérias na humanidade [7, 8]. O Projeto 1000 Genomas identificou um amplo espectro de variação genética, incluindo mais de 88 milhões de variantes, consistindo em 84,7 milhões de polimorfismos de nucleotídeo único (SNPs), 3,6 milhões de inserções/exclusões curtas (indels) e 60.000 variantes estruturais. O número total de variantes de nucleotídeos únicos (SNVs) no DNA mitocondrial acumuladas em humanos modernos é de 19.811, conforme relatado pelo MITOMAP.
Eventos Catastróficos como Indutores de Picos de Mutação
A radiação ionizante é um agente mutagênico conhecido que pode causar danos ao DNA, resultando em um aumento nas taxas de mutação [4][6]. Eventos como explosões nucleares, erupções vulcânicas e impactos de asteroides podem expor organismos a níveis elevados de radiação, levando a um acúmulo acelerado de mutações [5][7]. Além da radiação, outros estressores ambientais, como hipóxia severa, podem comprometer os sistemas de reparo do DNA [Lee et al., 2021][8].
A história do meteoro de Patterson (Cannyon diablo) que datou a terra em 4,5 bilhão de anos, menciona que Patterson falhou em hipotetizar possibilidades ligadas a efeitos de aceleração de elétrons na queda de asteroides, e complementamos que, totalmente provável que na historia deste meteoro , houve atuação de plasma, efeito piezoelétrico e altíssimas temperaturas que chamamos de EPPAT; todos aceleradores de elétrons, logicamente que tais efeitos aceleradores de elétrons (ver como funciona as tokamaks) , ultrapassariam a barreira de Coulomb e acelerariam o decaimento radioativo, explicando assim a aparência de rochas velhas quando contabilizamos decaimento constante atual dos elementos isótopos chumbo 207 e chumbo 206, bem como Rubídio – Estrôncio que ao datar 5,1 bilhão de anos, superficialmente confirmaria a datação em torno de 4,6 bilhão de anos que as pessoas comuns repetem (e até cientistas) sem atentar para este fato.
Esta situação compromete e anula totalmente a veracidade, chamada de absoluta, de todos os métodos de datação radiométrica de alta escala de tempo, e pode ser confirmada de diversas formas como no estudo abaixo onde se observa que há uma linha de tendência em relação a dizer que numericamente diâmetros maiores dos bólidos possuem maior probabilidade de estarem relacionados a maiores idades, e numericamente bólidos pequenos ajuntados em torno de idades menores, possuem proporcionalmente probabilidade de estarem relacionados a idades menores:


A admissão de uma chuva de asteroides (CA) na Terra desmantela o dogma da constância do decaimento radioativo, que fundamenta a datação radiométrica. Durante uma CA, uma combinação de temperatura, som, efeito piezoelétrico e plasma gerado por diferenciais de carga atua como aceleradores de elétrons, permitindo a superação da barreira de Coulomb e a perturbação de núcleos atômicos. Bólidos causariam aceleração de decaimento e derrubam o absolutismo datacional radiométrico, pois as experiências ligadas a fusão nuclear e aceleração de partículas , experimentadas principalmente em equipamentos do tipo Tokamak, quando associadas ao que se esperaria , em termos de efeitos aceleradores de partículas, na queda de grandes bólidos na terra; nos falam que ocorreu aceleração de decaimento radioativo em diversos pontos da terra , fazendo com que rochas passassem a ter a aparência de “envelhecidas”. A evidência de efeitos aceleradores de partículas durante impactos de asteroides, junto com a preservação de tecidos moles e a presença de carbono-14 em rochas muito antigas, destrói por completo a validade dos métodos tradicionais de datação geológica. O estudo examina as contradições que surgem ao confrontar a teoria da constância de decaimento com os efeitos de fenômenos físicos como o efeito piezoelétrico e a geração de plasma. A interação da Terra com corpos celestes, especialmente asteroides, moldou sua superfície e influenciou a vida. A análise de chuvas de asteroides é crucial para entender a geologia e e o aparecimento repentino de cenários geológicos e novas condições ambientais que fabricaram esse contraste do mundo fóssil , repetido morfologicamente sob paradoxo evolutivo e de estase de formas (Sodré , 2017), principalmente contraste quanto ao tamanho, longevidade, ausência de acúmulo mutacional e riqueza genética das espécies mães ancestrais . A integração de dados multidisciplinares é essencial para compreender os impactos e suas consequências.Este artigo questiona a validade universal da datação radiométrica, argumentando que os impactos de asteroides exerceram uma influência significativa nos processos geológicos e biológicos da Terra. A análise se baseia em evidências de catastrofismo, na potencial inconstância do decaimento radioativo e em anomalias encontradas em datações convencionais, sugerindo que eventos catastróficos podem ter acelerado a sedimentação, alterado a composição isotópica das rochas e promovido mudanças abruptas nas condições ambientais.
Recebi a contribuição de João Paulo Reis Braga citando que “Richard Milton, que mesmo não sendo defensor do movimento científico criacionismo da terra jovem, aponta, no entanto, que a prontidão em rejeitar datas radiométricas, exceto aquelas que fornecem “valores esperados”, é o motivo pelo qual vários métodos radiométricos podem ser considerados convergentes nas “eras” que “medem” (Milton, 1997, p. 49): “Assim, as datas publicadas sempre obedecem a datas preconcebidas e nunca as contradizem. Se todas as datas rejeitadas fossem recuperadas da cesta de lixo e adicionadas às datas publicadas, os resultados combinados mostrariam que as datas produzidas são a dispersão que se esperaria apenas pelo acaso” (Milton, 1997, p. 51) Milton, Richard. 1997. Shattering the myths of darwinism. Park Street Press, Rochester, VT.[41]
Em geral, as datas no ‘parque correto’ são consideradas corretas e são publicadas, mas aquelas em desacordo com outros dados raramente são publicadas… (Mauger, 1977, p. 37). In general, dates in the `correct ball park’ are assumed to be correct and are published, but those in disagreement with other data are seldom published…(Mauger, 1977, p. 37) MAUGER, Richard L. K-Ar ages of biotites from tuffs in Eocene rocks of the Green River, Washakiw and Uinta Basins[42].[43] Contributions to Geology, Wyoming University. 15(1):17, 1977.
“A aparente convergência de resultados de datação radiométrica é mais uma quimera do que realidade porque “muitas determinações de idade que não concordam com as escalas de tempo atualmente aceitas são simplesmente rejeitadas como erradas…” (Paul, 1980, p. 184) PAUL, Christopher RC. The natural history of fossils.[44] Holmes and Meier, New York, 1980.
Esse artigo em particular tem excelentes tabelas e referências bibliográficas mostrando discrepâncias nas datações[45]
A própria técnica radiométrica desbanca estas datações como podemos ver em:[46]
“Ossos de dinossauro foram datados por radiocarbono (carbono-14). As datas variam de 22.000 a 39.000 anos antes do presente”
The Data: Carbon-14 in dinosaur bones
| Dinosaur (a) | Lab/Method/Fraction (b,c,d) | C-14 Years BP (Before Present) | Date | USA State |
| Acro Acro Acro Acro Acro Allosaurus Hadrosaur #1 Hadrosaur #1 Triceratops #1 Triceratops #1 Triceratops #1 Triceratops #2 Triceratops #2 Hadrosaur #2 Hadrosaur #2 Hadrosaur #2 Hadrosaur #2 Hadrosaur #2 Hadrosaur #3 Apatosaur | GX-15155-A/Beta/bio GX-15155-A/AMS/bio AA-5786/AMS/bio-scrapings UGAMS-7509a/AMS/bio UGAMS-7509b/AMS/bow UGAMS-02947/AMS/bio KIA-5523/AMS/bow KIA-5523/AMS/hum GX-32372/AMS/col GX-32647/Beta/bow UGAMS-04973a/AMS/bio UGAMS-03228a/AMS/bio UGAMS-03228b/AMS/col GX-32739/Beta/ext GX-32678/AMS/w UGAMS-01935/AMS/bio UGAMS-01936/AMS/w UGAMS-01937/AMS/col UGAMS-9893/AMS/bio UGAMS-9891/AMS/bio | >32,400 25,750 + 280 23,760 + 270 29,690 + 90 30,640 + 90 31,360 + 100 31,050 + 230/-220 36,480 + 560/-530 30,890 + 200 33,830 + 2910/-1960 24,340 + 70 39,230 + 140 30,110 + 80 22,380 + 800 22,990 +130 25,670 + 220 25,170 + 230 23,170 + 170 37,660 + 160 38,250 + 160 | 11/10/1989 06/14/1990 10/23/1990 10/27/2010 10/27/2010 05/01/2008 10/01/1998 10/01/1998 08/25/2006 09/12/2006 10/29/2009 08/27/2008 08/27/2008 01/06/2007 04/04/2007 04/10/2007 04/10/2007 04/10/2007 11/29/2011 11/29/2011 | TX TX TX TX TX CO AK AK MT MT MT MT MT MT MT MT MT MT CO CO |
Marcos Nogueira Eberlin e sua Declaração Bombástica da Science ter Aprovado Artigos Classificados por ele como “porcarias”
Como mencionou Faymann “existem ciências e afirmações cientificas com maior ou menor graus de certeza”, na pesquisa de testes clínicos, um coordenador de testes clínicos de medicamentos ou dispositivos médicos, deve buscar testar, depois de prévios testes em laboratório e animais (pré-clínicos), em torno de 2000 casos em pacientes (fases clínicas 1, 2, 3 ) e por último, ainda sofre o teste do produto no mercado (fase 4), que em geral envolve centenas de milhares de pessoas, milhões ou até bilhões de pessoas. São muitos testes para poder se afirmar eficiência ou não de um dispositivo ou medicamento. Muito rigor científico e facilidade de testes , gera portanto um grau de certeza maior. Na paleontologia, não podemos seguir tal rigor, e por isso as vezes apenas um achado fóssil , pode gerar muitas interpretações e deduções; exatamente por isso a paleontologia se caracteriza como uma ciência mais interpretativa e conjectural , que factual. Quando influenciada por um modelo isto se torna ainda mais desapegada do rigor cientifico e submissa aquele modelo, a ponto do registro de fósseis de animais mortos ser interpretada como registro de surgimento de vida .
Muitas análises de engenharia de materiais, geologia, paleontologia, nutrição, medicina, farmácia, etc.. precisam da análise e do laudo químico; porque quando uma aárea de estudo depende da química ou bioquímica, ela gera autoridade para que o especialista naquela área decida a validade ou não de alguma afirmação.
Sobre as reações de Fenton publicadas por Mary Schwvatzer e outros[47][48], o bioquímico Dr. Marcos Eberlin, que tem 2 publicações analisando as reações de Fenton, classificou como “porcaria” tais publicações (incluindo da Science) que se escoram em reações de Fenton para preservação incrível de fosseis, onde Mary Schwatzer defendeu que óxido de ferro teria ajudado preservar tecido orgânico de T-rex por 68 milhões de anos baseada numa experiência , comparativamente fútil, de apenas 2 anos.
Muitos cientistas preferem questionar o “dogma” de que proteínas , DNA e biomoléculas não se preservam por muito tempo, do que questionar as “inerrantes” datações e “absolutos” métodos de datação, porque isso representa comprar uma imensa briga com a patrulha darwinista[49], e exatamente por isso, sabem que tais justificativas de tal impossível preservação orgânica em fósseis, serão aplaudidos e aceitos pelas revistas científicas. Precisamente é por este contexto de dominio do paradigma e recusa de ser implodido, que se consegue publicar milhares de artigos “científicos” categorizados por Eberlin e outros cientistas, como “porcarias” defendendo tal impossível possibilidade. Os mais militantes do darwinismo repetem ao mesmo tempo, que Eberlin , por nunca ter publicado artigo científico especificamente na área da polêmica design inteligente, ou criacionismo versus evolucionismo , ou paleontologia , ou evolução, não teria autoridade para opinar a respeito, por mais que análises bioquímicas pertençam ao universo do bioquímico e não universo da paleontologia e quaisquer outras áreas e polêmicas envolvidas.
Além dos testes químicos que estas áreas dependem gerar autoridade a Eberlin, devemos lembrar que ele é peeer review de diversas revistas em assuntos que ele já publicou, e demonstra ser especialista em muitos pontos bioquímicos dos quais depende outras áreas de estudo, além disso, as “reações de Eberlin” assim citadas em livros no mundo inteiro , reação de acetalização e transacetalização polar na fase gasosa, foram nomeadas em homenagem a ele, sendo cientista brasileiro [50][51], Scopus 73+5[52] e índice h 87[53] e citado na literatura científica 34.322 vezes, como ganhador da medalha Thomson de espectrometria de massas reconhecida pelos pares na área, em 2016. [54][55] e portanto pode sim declarar e julgar como porcaria publicações que envolvam reações bioquímicas como as de Fenton[56].
Novos modelos sobre a formação do manto terrestre tem sido propostos principalmente por equipes de geofísicos criacionistas ligados a John Baumgardner[1] que também questionou métodos absolutos, por meio de testes que contrastam idades atribuídas pela onipresença inesperada de carbono 14 (devido sua meia-vida curta) em materiais de origem orgânica incrustados em rochas consideradas antigas em torno de milhões e bilhões de anos[2][3]
A humanidade teve pico de acúmulo de genes deleterios entre 5 a 10.000 atrás e mais precisamente entre 2 e 6.000 anos atrás
Este artigo da Nature citado na tese de Crabtree sobre nosso frágil intelecto[152] e previsão de aumento exponencial de doenças neurológicas[125][126][130], nos mostra que houve inicio de acúmulo de genes deletérios entre 5 a 10.000 anos atrás, numa verdadeira explosão deles[38], como revela este estudo publicado[153]:
“Estudos em larga escala de variação genética humana relataram assinaturas de recente crescimento populacional explosivo, notáveis por um excesso de variantes genéticas raras, sugerindo que muitas mutações surgiram recentemente. Para avaliar quantitativamente mais a distribuição das idades de mutação, nós resequenciamos 15.336 genes em 6.515 indivíduos de ascendência americano e Africano Europeu e inferir a idade de 1.146.401 autossômicas variantes de nucleotídeo único (SNVS). Nós estimamos que cerca de 73% de todos os SNVs codificadores de proteínas e cerca de 86% de SNVs previsto para ser deletério surgiu nos últimos anos 5.000-10.000. A idade média dos SNVs deletérios variou significativamente entre vias moleculares e genes de doenças continha uma proporção significativamente maior de SNVs deletérios recentemente surgiram de outros genes. Além disso, os americanos europeus tiveram um excesso de variantes deletérias em genes essenciais e mendeliana doença em comparação com os afro-americanos, de acordo com fraca seleção purificadora, devido à dispersão Out-of-Africa”.
Temos hoje segundo banco de dados BLAST entre 15 a 88 milhões de mutações [7][8] com ” um amplo espectro de variação genética, no total, mais de 88 milhões de variantes (84,7 milhões de polimorfismos de nucleotídeo único (SNPs), 3,6 milhões de inserções/exclusões curtas (indels) e 60.000 variantes estruturais.” [154][155][156][157]em genes germinativos 100.000[158]. Se temos um acúmulo 150 mutações deletérias a cada 25 anos (geração), fica fácil mensurar quando aproximadamente tivemos pureza genética[159]. Um dado super interessante resumiu o Dr. Marcos Eberlin[160], unindo as taxas mutacionais e picos percebidos, que se acumulam geração após geração, e em seguida dividindo por geração em relação ao total de mutações identificadas no genoma humano[161][162] . Descobrimos que a apenas 6 a 12.000 anos, ou em torno de 10.000 anos[163] nós tínhamos pureza genética[164] , ou seja, isso confirma o relato bíblico arqueológico de Gênesis quando fala dos ancestrais iniciais Adão e Eva[165][166], bem como confirma genealogias estatísticas em torno de 6.000 anos como distância temporal dos patriarcas ancestrais da humanidade [167][168][12][169][170][171][172][173][174] sendo que, desde 2004, já se admitia que dos atuais vivos, “o MRCA (ancestral comum mais recente) de todos os humanos atuais viveu apenas alguns milhares de anos atrás.[175] e que vivos e mortos não poderia estar tão afastados.
O Contraste fóssil revela catástrofe que modificou o ambiente
A mudança drástica no ser vivo indica mudança drástica de ambiente[176][177][178]. Não temos gigantes sendo produzidos pela evolução hoje, hoje baleias e girafas estão em extinção, mas no registro fóssil eles são abundantes[179][180][181][182][183] . A mudança de ambiente pressiona os seres vivos a se adaptarem, variarem, e consequentemente empobrecerem geneticamente, uma destas mudanças pode estar ligada a atmosfera do planeta Terra, que detinha maior concentração de oxigênio o que favorecia ainda mais as formas de vida, longevidade , tamanho, e controle de patógenos como vírus, bactérias e fungos .. A oxigenação é fartamente citada na literatura como gerando múltiplos efeitos benéficos a saúde e diversas técnicas tem sido defendidas como ferramentas úteis nos tratamentos como câmaras hiperbáricas, ventiladores, balão de oxigênio e ozonioterapias[184]. O prefeito de Itajaí- SC, Brasil, médico, Dr. Volnei Morastoni, tem recomendado a aplicação retal de ozônio para pacientes que apresentem sintomas do novo coronavírus SARS-CoV-2 que manifesta Covid-19. Alguns ensaios clínicos tem sido publicados confirmando a eficiência desta técnica centenária para Covid-19[185] [186]. A técnica já conta mais de 3500 artigos no Pubmed e mais de 8000 artigos no Science Direct e desde a patente de Tesla em 1896 que se sabe dos múltiplos benefícios da ozonioterapia atuando no combate a 264 doenças incluindo efeitos antivirais, oxigenação, aspectos antinflamatórios e antidiabéticos[187][188][189], melhorando a circulação, combatendo hipertensão[190], grávidas hipertensas[191], doenças de pele[192] o que coloca a técnica como conversora de inúmeros benefícios conjuntos aos pacientes de risco, tantos, que ameaçam centenas de patentes de medicamentos, provocando perseguições de agencias do governo, e midia, muitas vezes controladas por lobbys da industria farmacêutica. Neste contexto dos benefícios do oxigênio, percebemos que a terra era ainda mais adaptável a vida , ainda mais bem projetada, e na sua falta, temos o aumento da entropia genética nas suas formas EGI e EGP.
Os haplogrupos mitocondriais M e N originaram-se na África subsaariana e espalharam-se pelo resto do mundo à medida que os primeiros humanos modernos migraram para fora da África há cerca de 60.000 anos (Behar et al., 2008). Esses haplogrupos ancestrais deram origem a grupos filhos, incluindo os haplogrupos L na África e os haplogrupos R e U na Eurásia. Os haplogrupos R e U espalharam-se ainda mais, dando origem aos haplogrupos H, J e T na Europa (Achilli et al., 2004).
O pico de mutações SNPs relatado por Fu et al. (2013) coincide temporalmente com a divergência desses haplogrupos europeus dos ancestrais M e N. Isso implica que hub por volta de 5.000 a 10.000 anos, o número de mutações transmitidas por linhagem materna aumentou acentuadamente na Eurásia, contribuindo para a diferenciação de haplogrupos como H, J e U.
Em conclusão, os dados genéticos de mais de 6.500 exomas humanos e de haplogrupos mitocondriais sugerem que ocorreu um aumento acentuado nas taxas de mutação genética materna na Eurásia em um período de 5.000 a 10.000 anos atrás. Este “pico” de mutações SNPs pode ter facilitado a divergência dos haplogrupos mitocondriais europeus H, J e U a partir de seus ancestrais africanos M e N. Mais pesquisas são necessárias para explorar as causas desse aumento repentino nas taxas de mutação e seu impacto na evolução humana recente.
Sobre o pico recente de mutações SNPs:
“Analisamos 6.515 exomas humanos da população geral e constatamos que a maioria das variantes de proteínas codificadas surgiu durante os últimos 5.000 a 10.000 anos” (Fu et al., 2013).
Sobre a origem e divergência dos haplogrupos mitocondriais:
“Os resultados indicam que as haplogrupos mitocondriais não-africanas M, N e R têm uma única origem no Estreito de Bering ou na Ásia Central há 50.000-65.000 anos e se diversificaram localmente na Eurásia” (Torroni et al., 2006).
“Os haplogrupos U4, U5, e U6 iniciaram separadamente do haplogrupo U, e expandiram-se logo após o último máximo glacial, há aproximadamente 8.000-15.000 anos atrás” (Pereira et al., 2012).
“Combinando estes resultados com aqueles de estudos recentes, podemos tentativamente traçar as grandes linhas do seguinte cenário para a expansão dos haplogrupos humanos mitocondriais modernos. As linhagens mitocondriais já conhecidas M, N e R surgiram na África subsaariana e no Oriente Médio antes da expansão humana inicial da Eurásia” (Behar et al., 2008).
” Os haplogrupos mitocondriais europeus mais difundidos, H, J, T, e U, originaram-se no Oriente Médio e no Cáucaso por volta de 30.000 anos atrás e expandiram-se na Europa após a última era glacial” (Achilli et al., 2004).
Baseado no contexto fornecido, uma combinação de fatores pode justificar o aumento significativo no número de mutações mitocondriais observadas entre as múmias e a população atual:
- Gargalo Populacional pelo Dilúvio Global:
- Um evento catastrófico como um dilúvio global provavelmente causou um severo gargalo populacional.
- Gargalos populacionais podem permitir a fixação de mutações deletérias que normalmente seriam eliminadas por seleção natural em populações maiores.
- Isso explicaria parte do aumento de 250 mutações observadas nas múmias para as 19.000 mutações atuais.
- Exposição à Radiação Ionizante:
- Além do gargalo populacional, um bombardeio intenso de radiação ionizante sobre os sobreviventes do dilúvio poderia ter acelerado significativamente a taxa de mutação mitocondrial.
- A radiação ionizante é um agente mutagênico conhecido que pode causar danos e mutações no DNA mitocondrial.
- Esse efeito sinérgico do gargalo populacional seguido de exposição à radiação poderia justificar o impressionante aumento de 75 vezes no número de mutações mitocondriais.
Portanto, a combinação de um severo gargalo populacional causado pelo dilúvio global, seguido de intensa exposição à radiação ionizante pelos sobreviventes, parece ser uma explicação plausível para o enorme acúmulo de mutações mitocondriais observado entre as múmias e a população atual. Essa hipótese está de acordo com os conceitos discutidos no documento fornecido sobre a dinâmica de mutações deletérias em populações sob perturbações demográficas.
Evidências de Picos de Mutação em Populações Antigas
Estudos de DNA antigo revelaram padrões de mutação que coincidem com períodos de estresse ambiental, sugerindo que eventos catastróficos influenciam a diversidade genética [9][9]. A análise de populações que sobreviveram a desastres naturais mostra um aumento nas taxas de mutação em comparação com populações que não foram expostas a tais eventos [10][10].
Mecanismos de Mutagênese Induzida por Catástrofes
O dano direto ao DNA por radiação e toxinas, junto com o estresse celular, pode resultar em um reparo de DNA prejudicado [11][11]. O impacto na fidelidade da replicação do DNA mitocondrial pode contribuir para a acumulação de mutações [12][12]. A exposição a radiações ionizantes superiores a 2 Gray resulta em uma deterioração significativa na atividade da PARP1, uma enzima crucial na detecção de lesões de DNA [Smith et al., 2022][13]. A hipóxia severa, frequentemente associada a eventos catastróficos, compromete significativamente os sistemas de reparo do DNA em níveis moleculares [Lee et al., 2021][14]. A radiação ionizante induz degradação proteolítica de sensores críticos como PARP1 e componentes do complexo MRN, comprometendo os mecanismos de reparo [Kim et al., 2020][15].
Implicações Evolutivas do Tipo Degenerativas
Picos de mutação podem atuar como um motor de rápida adaptação, onde mutações mitocondriais desempenham um papel chave na evolução humana [13][16]. No entanto, o acúmulo de mutações deletérias no genoma humano, conforme observado em vários bancos de dados genéticos, levanta preocupações sobre a degeneração genética. A compreensão desses mecanismos é crucial para a biologia evolutiva do tipo degenerativa e para a interpretação da diversidade genética nas populações modernas.
3 Ls matriarcais mitocondriais sob forte radiação e as oportunidades de pesquisa do câncer e longevidade
Evidências de um pico recente de mutações SNPs a divergência nítida e numerosa de SNPs em haplogrupos mitocondriais humanos e a separação violenta da crosta continental revelada pela alta radioatividade das rochas da borda da plataforma continental. Considerando a existência de 3 matriarcas descendentes da eva mitocondrial , com alta diferença entre si de mutações no mtDNA, concluimos que elas , sendo contemporâneas , para poderem ter tal diferenciação, participaram juntas de uma catastrofe envolvendo muita radioatividade, que seria praticamente a única hipótese provável para poder gerar em 3 mulheres contemporâneas e aparentadas, tal diferença de mutações mitocondriais entre si. Segundo Akey et e tal , há 5.115 anos iniciou o acúmulo de mutações gerais na humanidade (nuclear e mtDNA), ele justifica tal inicio de acúmulo devido o homem ter deixado de ser apenas caçador coletor e passado a ser agricultor , porém questionamos que tal justificativa seja insuficiente, pois mesmo supondo como fato, tal mudança de estilo de vida da humanidade para esta data, não haveria uma explosão de mutações deletérias quando o acúmulo delas ainda não existiam em 3.098 ac.
Perguntamos se a dedução de 3 matriarcas Ls mitocondriais sofrendo exposição a radiações, possa ter relação com as 3 noras de Noé, manu (hamanidade), pois são também apontadas por outras centenas de fontes arqueológicas (além da bíblica que é a mais conhecida) que teriam participado de uma catástrofe global acompanhada por um diluvio global na terra. Existe entre os vários modelos de diluvio criacionistas, um modelo que defende uma chuva de asteroides que teriam provocado diversas extinções dos tipos básicos ancestrais fósseis, que estão repetidos no registro fóssil (paradoxo da estase morfológica e pontualismo ou saltacionismo evolutivo pela ausência de graduação transicional) espelhando sepultamento de populações ancestrais fósseis. Diversas catástrofes associadas a diversas inundações globais geradas por transgressões e regressões marinhas nos continentes, provocando diversas formações geológicas , como as gigantescas erosões grossas e espessas margeando rios e córregos, que são as bacias sedimentares , onde seu material arrastado para o mar formariam as camadas sedimentares longas por milhares de kilômetros, espessas , segregadas em seus materiais fisico-químicos comuns, e largas, como as que vemos hoje, e sepultariam grande parte da vida marinha e aquática que possui representatividade no registro fóssil de 95% deles, enquanto animais vertebrados continentais possuem representação de apenas 0,01% no registro fóssil, o que combina perfeitamente com uma catástrofe envolvendo muita água carregando sedimentos para sepultar principalmente animais marinhos, que é para onde aguas catastróficas globais levaria a maior parte dos sedimentos.
Uma chuva de asteroides , em consonância com diversas simulações da NASA, provocaria diversos efeitos elétricos na terra produzindo velocidade gigantesca de elétrons, devido a conjugação de efeito piezoelétrico + atrito dos asteroides na crosta continental + diferencial de cargas+ som + altíssima temperatura imediata , gerando plasmas gigantes com altíssima velocidade de elétrons, devido a conjugação dos efeitos na queda dos mesmos. Tal modelo catastrófico, explicaria entre outros fenômenos , como a crosta continental de média de 70 km de espessura, pôde então ser violentamente fendida, rachada, cortada: Plasma é um dos melhores sistema de corte rápido e eficiente. Um plasma desta magnitude, geraria também uma velocidade de elétrons que ultrapassaria a barreira de coulomb, arrancando neutrons e prótons , principalmente das rochas por onde o plasma passou: as rochas de intersecção situadas nas bordas da plataforma continental. A pesquisa abaixo demonstrará evidências deste modelo, conjugando 3 noras de Noé, 3 Ls matriarcais mitocondrial, catastrofismo radioativo, queda brusca de tamanho em relação aos ancestrais fósseis, queda brusca de longevidade relatada nas genealogias pós diluvio, e neste ponto uma aplicação prática, porque esta relatada queda drástica de longevidade descrita na genealogia humana , que combina com altíssima radiação bombardeando DNA nuclear e mitocondrial nas noras de Noé, pode ter com muita probabilidade , altíssima relação com a destruição mutacional de sequências codificantes de proteínas reparadoras semelhantes a p53 p21 P 17; e isso pode gerar uma boa pesquisa no sentido de quem sabe recuperar essas sequências codificantes por meio de crisprcas9-12, e criar proteínas reparadoras que foram extintas, o que evitaria o câncer e diversas doenças geradas sobretudo pelo envelhecimento, bem como para prover sustentação para projetos de aumento de longevidade.
Um estudo de 2013 que analisou mais de 6.500 exomas humanos identificou que a maioria das variantes de proteínas codificadas (86%) surgiram nos últimos 5.000 a 10.000 anos (Fu et al., 2013). Este pico recente de mutações de nucleotídeo único (SNPs) coincide com a divergência dos três principais haplogrupos mitocondriais da Europa (haplogrupos H, J e U) dos haplogrupos ancestrais M e N, que surgiram na África (Torroni et al., 2006; Pereira et al., 2012).
Como poderia haver tantas mutações SNPs entre 5 e 10.000 anos atrás , poderia ser que 3 mulheres receberam uma dose exagerada de mutações numa catástrofe envolvendo muitas radiações? Descendendo seu mtDNA marcado desde então justificando porque haplogrupos mitocondriais se distingam tanto na genealogia? (lembrando que um pico de radiações faz com que ignoremos as datações aceitas, pois que as mesmas estão baseadas na constancia de decaimento e esta hipótese ao tentar explicar o pico de mutações recorre a inconstância de decaimento radioativo.
Essa é uma hipótese de um evento catastrófico envolvendo exposição a radiação poderia, em teoria, levar a um pico acentuado em mutações SNPs mitocondriais que foram então transmitidas para os descendentes dessas linhagens, nos leva a considerar alguns pontos:
-
Exposição aguda a radiação ionizante (como de uma explosão nuclear ou acidente envolvendo material radioativo) pode causar danos no DNA mitocondrial e induzir mutações SNPs. As mitocôndrias são particularmente sensíveis aos efeitos da radiação devido à proximidade do DNA mitocondrial às fontes de espécies reativas de oxigênio.
-
Se somente algumas linhagens maternas (por exemplo, as precursoras dos haplogrupos H, J e U) foram expostas a altos níveis de radiação, isso poderia explicar por que esses haplogrupos se tornaram tão distintos de M e N. Suas taxas de mutação teriam aumentado abruptamente, levando à acumulação de mutações SNPs.
-
Um evento catastrófico desse tipo poderia distorcer as estimativas de datação baseadas na taxa constante de mutações. Se as taxas de mutação aumentaram repentinamente devido à exposição à radiação, as linhagens afetadas pareceriam mais antigas do que realmente são.
Para ser plausível, precisaríamos identificar uma fonte possível de exposição aguda à radiação que tenha ocorrido na Eurásia há 5.000-10.000 anos e que possa ter afetado somente algumas linhagens maternas.
Altos Níveis de Radioatividade em Rochas da Borda da Plataforma Continental: Implicações para a Tese de Separação Continental Violenta
Revisamos estudos que documentam altos níveis de radioatividade em rochas da borda da plataforma continental, sugerindo que esses dados podem indicar processos geológicos violentos associados à separação continental. A análise de diferentes regiões geográficas revela a presença de radionuclídeos, como urânio, tório e potássio-40, em concentrações significativamente elevadas, o que pode fornecer evidências para a tese de uma separação continental extremamente violenta. Esta revisão expandida incorpora estudos adicionais para apoiar ainda mais a hipótese.
A radioatividade natural em rochas e solos é um indicador importante dos processos geológicos que moldaram a crosta terrestre[17]. A presença de radionuclídeos, como urânio e tório, está frequentemente associada a atividades magmáticas e tectônicas intensas[18]. Neste contexto, a análise de rochas da borda da plataforma continental pode oferecer insights sobre a dinâmica da separação continental[19]. Este artigo expande pesquisas anteriores, incluindo uma gama mais ampla de estudos e contextos geológicos. O foco está em demonstrar que a elevação da radioatividade é uma característica das margens continentais e que essa elevação pode estar ligada a eventos geológicos de alta energia[20].
Radioatividade Natural em Rochas e Solos
A radioatividade natural em rochas e solos é um fenômeno geológico que ocorre devido à presença de radionuclídeos como urânio, tório e potássio-40. Estudos têm demonstrado que a concentração desses elementos pode variar significativamente dependendo da localização geográfica e da composição geológica.
Estudos de Caso
-
Jordânia: Al-Hadid, M. et al. (2007) mediram a radioatividade em granitos usados na construção, encontrando níveis elevados de urânio, tório e potássio-40, sugerindo uma origem geológica com processos energéticos significativos.[21]
-
Índia: Rao, K. S. et al. (2013) analisaram rochas e areias de praia da barragem de Nagarjunasagar, encontrando altas concentrações de urânio e tório, possivelmente relacionadas a processos geológicos violentos na área.[22]
-
Antártica Oriental: Harris, D. et al. (2007) mediram a radioatividade em rochas crustais ricas em zircão, encontrando altos níveis de potássio-40, tório e urânio, indicando condições geológicas intensas.[23]
-
Brasil: Bittar, T. et al. (2012) revelaram altas concentrações de rádio em formações graníticas e gnaissicas no estado do Rio de Janeiro, sugerindo um passado tectônico violento.[24]
-
Turquia: Kurtulus, A. et al. (2012) encontraram níveis elevados de rádio, tório e urânio em rochas e areias de praia da região de Ezine, indicando que a área pode ter sido afetada por processos geológicos intensos.[25]
-
Pireneus: Pérez, J. et al. (1997) examinaram granitos dos Pireneus, encontrando altas concentrações de rádio e tório, sugerindo condições de alta energia durante a gênese do magma.[26]
-
Golfo de Bothnia: Kärkkäinen, N. et al. (2008) revelaram altas concentrações de elementos radioativos em granitos costeiros do nordeste do Golfo de Bothnia, implicando um processo magmático intenso na separação do Escudo Báltico-Uraliano.[27]
-
Arábia Saudita: Al-Saad, S. et al. (2001) encontraram altos teores de urânio e tório em granitoides do escudo árabe oriental, indicando uma formação tectônica violenta.[28]
-
Mar Morto: Ziv, B. et al. (2017) encontraram altos níveis de urânio e tório em solo ao longo do sistema de falhas de transformação do Mar Morto, sugerindo processos geológicos agressivos.[29]
-
Austrália: Williams, I. S. et al. (1992) identificaram desequilíbrio na série de urânio em granitos tectonicamente ativos da Lachlan Fold Belt, indicando mobilização violenta de elementos radioativos.[30]
Os resultados dos estudos revisados indicam que as rochas da borda da plataforma continental frequentemente exibem altos níveis de radioatividade, sugerindo que a separação continental foi marcada por processos geológicos intensos. A presença de radionuclídeos em concentrações elevadas pode ser indicativa de atividade magmática e tectônica violenta, corroborando a tese de que esses eventos foram cruciais na configuração atual da crosta terrestre. A elevação da radioatividade não é apenas uma ocorrência aleatória, mas está ligada a formações geológicas específicas e processos que são característicos de margens continentais e zonas de rift. A presença de desequilíbrio nas séries de urânio e tório apoia ainda mais a ideia de mobilização recente e de alta energia desses elementos.
A análise da radioatividade natural em rochas da borda da plataforma continental aponta para a ocorrência de eventos geológicos extremos associados à separação continental. O conjunto expandido de estudos reforça a ideia de que esses eventos foram violentos e energéticos. Estudos adicionais são necessários para aprofundar a compreensão dos mecanismos que geraram essas condições e suas implicações para a evolução da Terra. Especificamente, a pesquisa futura deve se concentrar na mapeação detalhada dos níveis de radioatividade ao longo das margens continentais, acompanhada de análises geoquímicas e geocronológicas para melhor restringir o tempo e a intensidade dos eventos tectônicos e magmáticos.
Impacto no mtDNA da radioatividade gerada no inicio da separação continental e rebaixamento de oxigênio na atmosfera
Para ser plausível, precisaríamos identificar uma fonte possível de exposição aguda à radiação que tenha ocorrido na Eurásia há 5.000-10.000 anos e que possa ter afetado somente algumas linhagens maternas.
A Mãe de Todos os Humanos: Eva Mitocondrial
O conceito de “Eva mitocondrial” – a ancestral matrilinear mais recente de todos os humanos vivos – tem sido objeto de extensos debates e pesquisas no campo das origens humanas. Enquanto alguns estudos propuseram uma linha do tempo de 200.000 anos ou mais para a Eva mitocondrial, uma análise mais detalhada das evidências disponíveis aponta para uma origem muito mais recente, provavelmente dentro dos últimos 6.000 anos. Aqui apresentamos 20 razões que apoiam uma Eva mitocondrial recente, juntamente com 20 razões que colocam em dúvida uma linha do tempo evolutiva profunda para as origens humanas.
20 Razões para Afirmar que a Eva Mitocondrial Viveu há 6.000 Anos
O baixo número de diferenças nucleotídeos entre os humanos modernos e a sequência “consenso da Eva”, com uma média de apenas 21,6 diferenças, é muito mais consistente com uma origem recente do que uma antiga.[31][32][33][34]
As taxas de mutação medidas no DNA mitocondrial humano sugerem uma idade muito mais jovem para a Eva mitocondrial, na ordem de milhares, e não de centenas de milhares de anos.
A falta de divergência genética significativa entre as populações humanas, todas sendo interfertéis, aponta para um ancestral comum recente, e não para divisões evolutivas profundas.
Modelagem matemática da diversidade do DNA mitocondrial demonstra que uma Eva mitocondrial vivendo há milhares, e não centenas de milhares de anos, se encaixa melhor nos dados.
A comparação das sequências de DNA mitocondrial de humanos e chimpanzés revela um nível surpreendentemente baixo de divergência, novamente sugerindo uma ancestralidade compartilhada recente.
As evidências arqueológicas e fósseis para a súbita aparição do comportamento e anatomia modernos humanos são mais compatíveis com uma origem recente do que um desenvolvimento evolutivo gradual ao longo de centenas de milênios.
Os relógios genéticos baseados nas taxas de mutação do DNA mitocondrial consistentemente produzem estimativas de idade para a Eva mitocondrial na faixa de milhares de anos, e não de centenas de milhares.
A endogamia e os gargalos populacionais observados na história humana, como os experimentados pelos neandertais e denisovanos, teriam sido letais sob um cenário de Eva mitocondrial antiga devido ao acúmulo de mutações prejudiciais, já que a maioria são deleterias e quase não existe estudos que comprovem alguma evolução pela atuação de mutações benéficas[35]
A falta de divergência genética significativa entre populações humanas é inexplicável sob um modelo de Eva mitocondrial antiga, uma vez que um desvio genético substancial deveria ter ocorrido ao longo de centenas de milhares de anos.
Análises de relógio molecular do DNA nuclear também apontam para uma origem relativamente recente para os humanos modernos, dentro dos últimos 10.000 anos, consistente com uma Eva mitocondrial na faixa de 6.000 anos atrás.
Considerando a diversidade do DNA mitocondrial observada em outras espécies, como chimpanzés, sugere que a diversidade mitocondrial humana é anormalmente baixa, mais alinhada com um ancestral comum recente.
A súbita aparição da tecnologia e cultura humanas modernas, como o surgimento da agricultura, escrita e estruturas sociais complexas, é melhor explicada por um evento de criação recente do que por um desenvolvimento evolutivo gradual ao longo de centenas de milhares de anos.
O padrão consistente da diversidade genética humana, com todas as populações sendo mais próximas entre si do que dos chimpanzés, é mais facilmente explicado por uma ancestralidade compartilhada recente do que por divisões evolutivas profundas.
A análise comparativa do DNA mitocondrial de restos humanos antigos, como os neandertais e denisovanos, indica que essas linhagens humanas arcaicas divergiram da linhagem humana moderna relativamente recentemente, dentro dos últimos 50.000 anos.
A falta de mistura genética significativa entre humanos modernos e outras espécies humanas arcaicas, como neandertais e denisovanos, é mais consistente com uma ancestralidade comum recente do que uma profunda divergência evolutiva.
As análises filogenéticas do DNA mitocondrial humano consistentemente colocam a Eva mitocondrial na raiz da árvore genealógica humana, com todas as linhagens humanas modernas descendendo dela, apoiando uma origem recente.
Os padrões biogeográficos da diversidade genética humana, com a maior diversidade observada na África subsaariana, são mais facilmente explicados por uma origem recente naquela região do que por uma origem africana antiga seguida de migrações.
A observação de altos níveis de diversidade genética dentro de certas populações humanas, como grupos indígenas na África, é mais consistente com um ancestral comum recente do que com uma divergência antiga.
A aparente falta de diferenças genéticas significativas entre populações humanas, apesar de sua separação geográfica e diversidade cultural, aponta para uma ancestralidade compartilhada recente e não para divisões evolutivas profundas.
O relato bíblico das origens humanas, que coloca a criação dos primeiros humanos, Adão e Eva, aproximadamente há 6.000 anos, é consistente com as evidências de uma Eva mitocondrial recente.
20 Razões que Colocam em Dúvida uma Eva Mitocondrial de 200.000 Anos
O baixo número de diferenças nucleotídicas entre os humanos modernos e a sequência “consenso da Eva”, com uma média de apenas 21,6 diferenças, é muito baixo para ter se acumulado ao longo de centenas de milhares de anos, dada a taxa estimada de mutação do DNA mitocondrial.
A diversidade genética dentro das populações humanas é notavelmente baixa em comparação com outras espécies, o que coloca em dúvida a ideia de que os humanos descendem de uma população que existiu há centenas de milhares de anos.
A súbita aparição do comportamento e tecnologia modernos humanos, como o uso complexo de ferramentas, arte e o desenvolvimento da agricultura, é difícil de reconciliar com uma Eva mitocondrial de 200.000 anos.
A falta de divergência genética significativa entre as populações humanas, todas sendo interfertéis, é incompatível com uma Eva mitocondrial antiga, pois um desvio genético substancial deveria ter ocorrido ao longo de centenas de milhares de anos.
Análises de relógio molecular de DNA mitocondrial e nuclear consistentemente produzem estimativas de idade para a Eva mitocondrial que são muito mais jovens do que 200.000 anos.
A endogamia e os gargalos populacionais observados na história humana, como os experimentados pelos neandertais e denisovanos, teriam sido letais sob um cenário de Eva mitocondrial antiga devido ao acúmulo de mutações prejudiciais.
A súbita aparição da anatomia e fisiologia humanas modernas, sem formas de transição claras no registro fóssil, é mais consistente com um evento de criação recente do que com um desenvolvimento evolutivo gradual ao longo de centenas de milênios.
A comparação das sequências de DNA mitocondrial de humanos e chimpanzés revela um nível surpreendentemente baixo de divergência, colocando em dúvida a ideia de que humanos e chimpanzés compartilharam um ancestral comum há centenas de milhares de anos.
O padrão consistente da diversidade genética humana, com todas as populações sendo mais próximas entre si do que aos chimpanzés, é difícil de reconciliar com um cenário de Eva mitocondrial antiga.
As análises filogenéticas do DNA mitocondrial humano consistentemente colocam a Eva mitocondrial na raiz da árvore genealógica humana, com todas as linhagens humanas modernas descendendo dela, apoiando uma origem recente.
Os padrões biogeográficos da diversidade genética humana, com a maior diversidade observada na África subsaariana, são mais facilmente explicados por uma origem recente naquela região do que por uma origem africana antiga seguida de migrações.
A observação de altos níveis de diversidade genética dentro de certas populações humanas, como grupos indígenas na África, é mais consistente com um ancestral comum recente do que com uma divergência antiga.
A aparente falta de diferenças genéticas significativas entre populações humanas, apesar de sua separação geográfica e diversidade cultural, aponta para uma ancestralidade compartilhada recente e não para divisões evolutivas profundas.
O relato bíblico das origens humanas, que coloca a criação dos primeiros humanos, Adão e Eva, aproximadamente há 6.000 anos, é consistente com as evidências de uma Eva mitocondrial recente.
A súbita emergência de uma linguagem humana complexa, que parece ter ocorrido relativamente recentemente em nossa história evolutiva, é difícil de reconciliar com um cenário de Eva mitocondrial antiga.
A falta de mistura genética significativa entre humanos modernos e outras espécies humanas arcaicas, como neandertais e denisovanos, é mais consistente com uma ancestralidade comum recente do que uma profunda divergência evolutiva.
A análise coeve mithocodrial wiki codigomparativa do DNA mitocondrial de restos humanos antigos, como os neandertais e denisovanos, indica que essas linhagens humanas arcaicas divergiram da linhagem humana moderna relativamente recentemente, dentro dos últimos 50.000 anos.
O nível notavelmente baixo de diversidade genética observado na espécie humana, especialmente em comparação com outros mamíferos, é incompatível com um cenário de Eva mitocondrial antiga, pois um desvio genético substancial deveria ter ocorrido ao longo de centenas de milhares de anos.
O padrão consistente da diversidade genética humana, com todas as populações sendo mais próximas entre si do que aos chimpanzés, é difícil de reconciliar com um cenário de Eva mitocondrial antiga que envolveria divisões evolutivas profundas.
A súbita aparição da tecnologia e cultura humanas modernas, como o surgimento da agricultura, escrita e estruturas sociais complexas, é melhor explicada por um evento de criação recente do que por um desenvolvimento evolutivo gradual ao longo de centenas de milhares de anos.
Haplogrupos
Os haplogrupos mitocondriais M e N originaram-se na África subsaariana e espalharam-se pelo resto do mundo à medida que os primeiros humanos modernos migraram para fora da África há cerca de 60.000 anos (Behar et al., 2008). Esses haplogrupos ancestrais deram origem a grupos filhos, incluindo os haplogrupos L na África e os haplogrupos R e U na Eurásia. Os haplogrupos R e U espalharam-se ainda mais, dando origem aos haplogrupos H, J e T na Europa (Achilli et al., 2004).
O pico de mutações SNPs relatado por Fu et al. (2013) coincide temporalmente com a divergência desses haplogrupos europeus dos ancestrais M e N. Isso implica que hub por volta de 5.000 a 10.000 anos, o número de mutações transmitidas por linhagem materna aumentou acentuadamente na Eurásia, contribuindo para a diferenciação de haplogrupos como H, J e U.
Em conclusão, os dados genéticos de mais de 6.500 exomas humanos e de haplogrupos mitocondriais sugerem que ocorreu um aumento acentuado nas taxas de mutação genética materna na Eurásia em um período de 5.000 a 10.000 anos atrás. Este “pico” de mutações SNPs pode ter facilitado a divergência dos haplogrupos mitocondriais europeus H, J e U a partir de seus ancestrais africanos M e N. Mais pesquisas são necessárias para explorar as causas desse aumento repentino nas taxas de mutação e seu impacto na evolução humana recente.
Sobre o pico recente de mutações SNPs:
“Analisamos 6.515 exomas humanos da população geral e constatamos que a maioria das variantes de proteínas codificadas surgiu durante os últimos 5.000 a 10.000 anos” (Fu et al., 2013).
Sobre a origem e divergência dos haplogrupos mitocondriais:
“Os resultados indicam que as haplogrupos mitocondriais não-africanas M, N e R têm uma única origem no Estreito de Bering ou na Ásia Central há 50.000-65.000 anos e se diversificaram localmente na Eurásia” (Torroni et al., 2006).
“Os haplogrupos U4, U5, e U6 iniciaram separadamente do haplogrupo U, e expandiram-se logo após o último máximo glacial, há aproximadamente 8.000-15.000 anos atrás” (Pereira et al., 2012).
“Combinando estes resultados com aqueles de estudos recentes, podemos tentativamente traçar as grandes linhas do seguinte cenário para a expansão dos haplogrupos humanos mitocondriais modernos. As linhagens mitocondriais já conhecidas M, N e R surgiram na África subsaariana e no Oriente Médio antes da expansão humana inicial da Eurásia” (Behar et al., 2008).
” Os haplogrupos mitocondriais europeus mais difundidos, H, J, T, e U, originaram-se no Oriente Médio e no Cáucaso por volta de 30.000 anos atrás e expandiram-se na Europa após a última era glacial” (Achilli et al., 2004).
Crispr cas9 e Sistemas de Edição Genética
CRISPR (repetições palindrômicas curtas regularmente interespaçadas agrupadas) refere-se a um sistema de defesa imune bacteriana que pode ser reaproveitado para edição de genoma direcionado . O sistema CRISPR-Cas9, especialmente, fornece um método rápido, barato e extremamente preciso de alterar sequências de DNA[36][37]. Os pesquisadores já usaram o CRISPR-Cas9 em modelos pré-clínicos para corrigir mutações que causam distrofia muscular de Duchenne [38], fibrose cística e vários outros distúrbios genéticos[39].
Outros sistemas CRISPR também estão sendo aproveitados inclusive sistemas de edição de mtDNA demonstram ser promissores [40]. CRISPR-Cas12 pode editar transcrições de RNA , enquanto CRISPR-Cas13 permite a manipulação de vírus de RNA como influenza e zika[41] . Juntas, essas ferramentas versáteis prometem a capacidade de editar com precisão as mutações causadoras de doenças do genoma humano com acúmulo em torno de 100 milhões delas[42]. A edição do gene CRISPR poderia, assim, retardar ou mesmo reverter o aumento exponencial de doenças genéticas hereditárias .
Perceber esse potencial ao abordar questões éticas exigirá diretrizes e políticas criteriosas para a tradução clínica. Mas o poder das tecnologias de grande sucesso CRISPR dá motivos genuínos para otimismo sobre o alívio da enorme carga humana de doenças genéticas nas próximas décadas.
Pesquisas apontam que estas doenças consideradas DCTN, que não são transmissíveis diretamente , podem ser passadas aos descendentes, pois uma mutação já presente ou adquirida, ou no esperma produzido[43][44], gerada por estilo de vida e aspectos epigenéticos, vírus, radiações e poluições, ou um radical livre gerado por má alimentação e/ou um metal pesado agrotóxico gerando mutações, antes ou durante a fase de reprodução, poderá acarretar em defeitos genéticos á prole. Estas pesquisas revelam aspectos extremos e preocupantes para aumento de prevalência relacionada a diabetes e hipertensão, onde o sono reparador de DNA da célula, torna-se ainda mais necessário para a diminuição de frequência e acúmulo nas descendências.[45]
A entropia geral de todas as coisas apontam um passado cada vez mais próximo de informações genéticas cada vez mais bem planejadas e menos afetadas pelo acúmulo contínuo de mutações deletérias, e neste aspecto estudos genéticos pretéritos comparados aos atuais demonstram ser uma ferramenta importante para o bom diagnóstico bem como quais as falhas deve receber maior prioridade seja pela medicina gênica, CRISPR cas9, ou pelas compensações proteômicas (suplementos alimentares) mais importantes em relação a constante degeneração humana, sendo as proteínas do cérebro um dos alvos mais relevantes uma vez que neurônios são afetados por atacado (em média 120 relações neurais são prejudicadas em função de apenas uma mutação[46][47])”um relaxamento da seleção contra mutações levemente deletérias, incluindo aquelas que aumentam a própria taxa de mutação. A consequência de longo prazo de tais efeitos é uma deterioração genética esperada na condição humana básica, potencialmente mensurável na escala de tempo de algumas gerações nas sociedades ocidentalizadas, e porque o cérebro é um alvo mutacional particularmente grande, isso é de particular preocupação”[48].
A humanidade teve pico de acúmulo de genes deletérios entre 5 a 10.000 atrás e mais precisamente entre 2 e 6.000 anos atrás
Este artigo da Nature citado na tese de Crabtree sobre nosso frágil intelecto[49] e previsão de aumento exponencial de doenças neurológicas[50][51][52], nos mostra que houve inicio de acúmulo de genes deletérios entre 5 a 10.000 anos atrás, numa verdadeira explosão deles[53], como revela este estudo publicado[54]:
“Estudos em larga escala de variação genética humana relataram assinaturas de recente crescimento populacional explosivo, notáveis por um excesso de variantes genéticas raras, sugerindo que muitas mutações surgiram recentemente. Para avaliar quantitativamente mais a distribuição das idades de mutação, nós resequenciamos 15.336 genes em 6.515 indivíduos de ascendência americano e Africano Europeu e inferir a idade de 1.146.401 autossômicas variantes de nucleotídeo único (SNVS). Nós estimamos que cerca de 73% de todos os SNVs codificadores de proteínas e cerca de 86% de SNVs previsto para ser deletério surgiu nos últimos anos 5.000-10.000. A idade média dos SNVs deletérios variou significativamente entre vias moleculares e genes de doenças continha uma proporção significativamente maior de SNVs deletérios recentemente surgiram de outros genes. Além disso, os americanos europeus tiveram um excesso de variantes deletérias em genes essenciais e mendeliana doença em comparação com os afro-americanos, de acordo com fraca seleção purificadora, devido à dispersão Out-of-Africa”.
Temos hoje segundo banco de dados BLAST entre 15 a 88 milhões de mutações [42][55] com ” um amplo espectro de variação genética, no total, mais de 88 milhões de variantes (84,7 milhões de polimorfismos de nucleotídeo único (SNPs), 3,6 milhões de inserções/exclusões curtas (indels) e 60.000 variantes estruturais.” [56][57][58][59]em genes germinativos 100.000[60]. Se temos um acúmulo 150 mutações deletérias a cada 25 anos (geração), fica fácil mensurar quando aproximadamente tivemos pureza genética[61]. Um dado super interessante resumiu o Dr. Marcos Eberlin[62], unindo as taxas mutacionais e picos percebidos, que se acumulam geração após geração, e em seguida dividindo por geração em relação ao total de mutações identificadas no genoma humano[63][64] . Descobrimos que a apenas 6 a 12.000 anos, ou em torno de 10.000 anos[65] nós tínhamos pureza genética[66] , ou seja, isso confirma o relato bíblico arqueológico de Gênesis quando fala dos ancestrais iniciais Adão e Eva[67][68], bem como confirma genealogias estatísticas em torno de 6.000 anos como distância temporal dos patriarcas ancestrais da humanidade [69][70][71][72][73][74][75][76][77] sendo que, desde 2004, já se admitia que dos atuais vivos, “o MRCA (ancestral comum mais recente) de todos os humanos atuais viveu apenas alguns milhares de anos atrás.[78] e que vivos e mortos não poderia estar tão afastados.
O Contraste fóssil revela catástrofe que modificou o ambiente
A mudança drástica no ser vivo indica mudança drástica de ambiente[79][80][81]. Não temos gigantes sendo produzidos pela evolução hoje, hoje baleias e girafas estão em extinção, mas no registro fóssil eles são abundantes[82][83][84][85][86] . A mudança de ambiente pressiona os seres vivos a se adaptarem, variarem, e consequentemente empobrecerem geneticamente, uma destas mudanças pode estar ligada a atmosfera do planeta Terra, que detinha maior concentração de oxigênio o que favorecia ainda mais as formas de vida, longevidade , tamanho, e controle de patógenos como vírus, bactérias e fungos .. A oxigenação é fartamente citada na literatura como gerando múltiplos efeitos benéficos a saúde e diversas técnicas tem sido defendidas como ferramentas úteis nos tratamentos como câmaras hiperbáricas, ventiladores, balão de oxigênio e ozonioterapias[87]. O prefeito de Itajaí- SC, Brasil, médico, Dr. Volnei Morastoni, tem recomendado a aplicação retal de ozônio para pacientes que apresentem sintomas do novo coronavírus SARS-CoV-2 que manifesta Covid-19. Alguns ensaios clínicos tem sido publicados confirmando a eficiência desta técnica centenária para Covid-19[88] [89]. A técnica já conta mais de 3500 artigos no Pubmed e mais de 8000 artigos no Science Direct e desde a patente de Tesla em 1896 que se sabe dos múltiplos benefícios da ozonioterapia atuando no combate a 264 doenças incluindo efeitos antivirais, oxigenação, aspectos antinflamatórios e antidiabéticos[90][91][92], melhorando a circulação, combatendo hipertensão[93], grávidas hipertensas[94], doenças de pele[95] o que coloca a técnica como conversora de inúmeros benefícios conjuntos aos pacientes de risco, tantos, que ameaçam centenas de patentes de medicamentos, provocando perseguições de agencias do governo, e midia, muitas vezes controladas por lobbys da industria farmacêutica. Neste contexto dos benefícios do oxigênio, percebemos que a terra era ainda mais adaptável a vida , ainda mais bem projetada, e na sua falta, temos o aumento da entropia genética nas suas formas EGI e EGP.
Os haplogrupos mitocondriais M e N originaram-se na África subsaariana e espalharam-se pelo resto do mundo à medida que os primeiros humanos modernos migraram para fora da África há cerca de 60.000 anos (Behar et al., 2008). Esses haplogrupos ancestrais deram origem a grupos filhos, incluindo os haplogrupos L na África e os haplogrupos R e U na Eurásia. Os haplogrupos R e U espalharam-se ainda mais, dando origem aos haplogrupos H, J e T na Europa (Achilli et al., 2004).
O pico de mutações SNPs relatado por Fu et al. (2013) coincide temporalmente com a divergência desses haplogrupos europeus dos ancestrais M e N. Isso implica que hub por volta de 5.000 a 10.000 anos, o número de mutações transmitidas por linhagem materna aumentou acentuadamente na Eurásia, contribuindo para a diferenciação de haplogrupos como H, J e U.
Em conclusão, os dados genéticos de mais de 6.500 exomas humanos e de haplogrupos mitocondriais sugerem que ocorreu um aumento acentuado nas taxas de mutação genética materna na Eurásia em um período de 5.000 a 10.000 anos atrás. Este “pico” de mutações SNPs pode ter facilitado a divergência dos haplogrupos mitocondriais europeus H, J e U a partir de seus ancestrais africanos M e N. Mais pesquisas são necessárias para explorar as causas desse aumento repentino nas taxas de mutação e seu impacto na evolução humana recente.
Sobre o pico recente de mutações SNPs:
“Analisamos 6.515 exomas humanos da população geral e constatamos que a maioria das variantes de proteínas codificadas surgiu durante os últimos 5.000 a 10.000 anos” (Fu et al., 2013).
Sobre a origem e divergência dos haplogrupos mitocondriais:
“Os resultados indicam que as haplogrupos mitocondriais não-africanas M, N e R têm uma única origem no Estreito de Bering ou na Ásia Central há 50.000-65.000 anos e se diversificaram localmente na Eurásia” (Torroni et al., 2006).
“Os haplogrupos U4, U5, e U6 iniciaram separadamente do haplogrupo U, e expandiram-se logo após o último máximo glacial, há aproximadamente 8.000-15.000 anos atrás” (Pereira et al., 2012).
“Combinando estes resultados com aqueles de estudos recentes, podemos tentativamente traçar as grandes linhas do seguinte cenário para a expansão dos haplogrupos humanos mitocondriais modernos. As linhagens mitocondriais já conhecidas M, N e R surgiram na África subsaariana e no Oriente Médio antes da expansão humana inicial da Eurásia” (Behar et al., 2008).
” Os haplogrupos mitocondriais europeus mais difundidos, H, J, T, e U, originaram-se no Oriente Médio e no Cáucaso por volta de 30.000 anos atrás e expandiram-se na Europa após a última era glacial” (Achilli et al., 2004).
Plantas Selvagens degeneradas necessitadas de melhoramento genético
As plantas cruzam entre espécies, gêneros, até o nível de família em enxertos[96][97][98][99] . Passando este limite não costuma dar nada demonstrando que o nível em torno de família se finda as possibilidades de cruzamentos artificiais diretos e indiretos, e se define também o limite da ancestralidade comum da proposta evolucionista extrapolada, de ancestralidade totalmente comum; restrito agora por este critério em torno do táxon familia, circunscrito ao cenário taxonômico monofilético e não parafilético como já criticam os taxonomistas: [100][101][102]
“Como lembra Rieppel (2005), o ramo das ciências biológicas chamado sistemática alcançou hoje maturidade justamente porque pretende substituir grupos não-monofiléticos por grupos monofiléticos, conforme preconizou Hennig (1966). A cladística permite que jantemos dinossauros tranqüilamente, na segurança das nossas casas, sem que para isso tenhamos que entrar em uma máquina miraculosa e viajar para o passado.”…”Essas hipóteses não podem ser confrontadas à luz de novas evidências ou a partir da análise de sua coerência interna: classificações da taxonomia clássica não são científicas, visto que não configuram hipóteses testáveis ou falseáveis (cf. Popper, 1959, 1962, 1972). As chamadas árvores evolutivas da taxonomia clássica são apenas asserções sem fundamentação metodológica adequada.”
Estes limites taxonômicos de ancestralidade em torno do táxon familia também lembra o que escreveu o bioquímico Michael Behe quando apontou “o limite do evolucionismo” extrapolado darwinista.[103][104] ao falar da complexidade irredutível de sistemas que precisam estar com milhares de mecanismos prontos de uma só vez pra poder funcionar/viver, [105] a começar do elemento mais básico, os aminoácidos, “que não se ligam sozinhos” como afirmou talvez o mais relevante químico sintético de todos os tempos, o Dr.James Tour[106]
Plantas Gigantes no Registro Fóssil
Ao olharmos a paleontologia percebemos confirmando o EGP no padrão gigante das “plantas originais”[107] [108]ou criadas ( que os biólogos criacionistas citados por Darwin tanto deduziam) , tipos básicos ancestrais em geral gigantes , estes que surgiram “prontas” no registro fóssil conforme os milhares de paleontólogos pontualistas encabeçados por Gould & Eldredge; e que o especialista em plantas Dr. Lonning[109], milhares de cientistas do Design Inteligente (que em 2004 compreendiam 11% dos cientistas), baraminology e criacionistas, tanto alegam que só podem ter sido criados já que o ser vivo mais simples precisaria de no mínimo umas 300.000 complexidades acontecendo de uma vez (Behe) pra poder viver .[110][111][112][113][114][115][116]
Estas gloriosas espécies tataravós descenderam sob taxas de acúmulo de mutações 99,9..⁹% deletérias, não filtradas pela peneira grossa da seleção natural[117] nossas espécies selvagens de hoje, e se não bastasse o acúmulo de mutações e defeitos causados por radiação atômica, incluindo o carbono 14 que passaram a estar presentes em todos os organismos, inserção de vírus degenerados[118] , se degeneraram ainda mais pelo próprio mecanismo de seleção natural o qual elimina parte do pool gênico, empobrecendo a já degenerada ao eliminar genes da população não selecionada( Marcos Ariel ) “esses aspectos da seleção natural, que geralmente levam ao empobrecimento do pool genético”[119]. E pensa que acabou? Não. Se degeneraram também pelas gambiarras (também classificadas como “alta sofisticação”) que a adaptação sempre exige e a EGP ocorre mais nitidamente pela deriva genética que obviamente divide pool gênico a cada deriva gerada por algum tipo de segregação, isolamentos , acidentes, mortandade.
Adolph Hitler idolatrava a ciência de sua época encabeçada inclusive pelo primo de Darwin, o Dr. Francis Galton (1822-1911) da eugenia, que inclusive acertou em cheio advertindo Darwin que ele não devesse casar com sua prima para que não tivesse uma prole adoecida, como de fato ocorreu, mas Hitler achava como muitos acham hoje, que a ciência daquela época era absolutamente correta em função de alguns acertos , considerava como sendo “o caminho a verdade e a vida ” , e por causa disto se dispôs a salvar a humanidade por meio da seleção nazista artificial da raça pura dos arianos, mas como toda ciência é apenas uma afirmação provisória, ele acabou por empobrecer a humanidade ao selecionar alguns e eliminar muitos judeus. Ainda bem que sobreviveram muitos judeus entre nós , se bem que tem alguns judeus que não gostam de cruzar com gentios (não judeus) porque a única coisa que não tem limite, Darwin deveria ter atentado a isso , é a crença desenfreada que de tempos em tempos toma conta da ciência em seu coletivismo extremamente falho e humano.
Naturalmente em geral todas as espécies empobrecem
Resultado da EGP? Nossas degeneradas e anãs espécies vegetais selvagens , as quais que não saciariam a fome da humanidade , nem dos pets com o mirrado milho selvagem, nem nossas crianças com frutos selvagens e tantas outras espécies castigadas pela EGP, que só ganham volume e características com propósito alimentício , quando se tenta artificialmente cruzar e re-ajuntar os genes derivados, selecionados, mutados; traduzindo em uma realidade decepcionadora daqueles que acreditavam que Darwin nos daria X-man em milhões de anos, quando se percebe que de fato acabou nos dando Hug Jakmans com câncer de pele.
Hoje, para o agricultor resgatar um pouquinho das plantas antigas, como uma samambaia gigante ou as gigantescas raízes fósseis gigantes das camadas sedimentares[120] carboníferas[121][122], temos que ajuntar o que sobrou dos genes derivados , dos não eliminados ou selecionados , os não mutados, evitando ao máximo os genes gambiarrados, por meio de infinitos cruzamentos ( melhoramento genético) . Também precisaremos usar a famosa biobalistica ( inventada pelo ex ateu e ex evolucionista John C Sanford[123][124][125][126][127][128]associados que hoje é criacionista terra jovem ) pra quem sabe, poder inserir a força, literalmente na bala, um gene ou outro que não conseguiríamos cruzar por causa dos limites e descontinuidade entre grupos de espécies[129][130][131], já que os limites de cruzamentos ou o ajuntamento do que sobrou não é suficiente , até quem sabe obter, depois de muito trabalho uma espécie melhorada, de genes re-ajuntados, para recapitular a historia da degeneração das espécies (TDE) e chegar pelo menos um pouco mais perto da gloriosa tataravó fóssil.
Depois de todo esforço de melhoramento artificial , o agricultor deve vigiar ao máximo sua planta protegendo-a de polinizar e de cruzar com plantas selvagens para não haja aquele milho bonito da Monsanto, Embrapa, Agroeste Sementes, CanaVialis, Alellyx, Cargill, Nestlé e Wal-Mart e outras .
Todas as plantas estão em geral degeneradas , ao estudarmos a quantidade de mutações e as taxas delas que se acumulam , concluímos que viemos de um paraíso genético , e devemos agradecer a técnicas de melhoramento genético e a cientistas como John C. Sanford, que é inventor da técnica geneGun de melhoramento genético, para que a população humana sobreviva com custos sustentáveis em sua alimentação.
Conclusão
Os dados sugerem que um pico de mutações durante catástrofe em torno de 5115 anos atrás, é uma explicação viável para a discrepância nas taxas de acúmulo de mutações mitocondriais histórica versus atuais. A compreensão desses mecanismos é crucial para a biologia evolutiva e para a interpretação da diversidade genética nas populações modernas, bem como para entender como reverter a perda de longevidade e aumento exponencial de doenças na humanidade ao editar genes mutados e imitar genes de nossos ancestrais, recuperando inclusive sistemas de reparo. As evidências apresentadas aqui sugerem fortemente que a Eva mitocondrial viveu muito mais recentemente do que a linha do tempo de 200.000 anos proposta por alguns estudos. A baixa diversidade genética observada nos humanos modernos, a súbita emergência do comportamento e tecnologia humanos modernos e o padrão consistente das relações genéticas apontam para uma Eva mitocondrial vivendo dentro dos últimos 6.000 anos, em linha com o relato bíblico das origens humanas. Embora o momento preciso ainda possa ser debatido, as evidências acumuladas apoiam de forma esmagadora uma origem recente para a Eva mitocondrial e para a espécie humana como um todo.
Referências
- Eyre-Walker, A. & Keightley, P. D. The distribution of fitness effects of new mutations. Nat. Rev. Genet. 8, 610–618 (2007).
- Brown, W. M., Prager, E. M., Wang, A. & Wilson, A. C. Mitochondrial DNA sequences of primates: Tempo and mode of evolution. J. Mol. Evol. 4, 225–239 (1982). doi:10.1007/bf01734101.
- ConfirmedMutations < MITOMAP < Foswiki. MITOMAP. Disponível em: www.mitomap.org. (Acesso em: 20 mar. 2025).
- Brown, W. M., Prager, E. M., Wang, A. & Wilson, A. C. Mitochondrial DNA sequences of primates: Tempo and mode of evolution. J. Mol. Evol. 4, 225–239 (1982). doi:10.1007/bf01734101.
- Coia, V. Mitochondrial diversity of modern Sudanese populations: Genetic contributions of sub-Saharan and North African populations. Hum. Biol. 88, 203–214 (2016).
- Howell, N. Evolutionary conservation of protein regions in the protonmotive cytochrome b gene and their possible roles in redox catalysis. J. Mol. Evol. 33, 157–167 (1991).
- Parsons, T. J. A high observed substitution rate in the human mitochondrial DNA control region. Nat. Genet. 15, 363–368 (1997).
- Lee, J. H. Severe hypoxia significantly impairs DNA repair systems at the molecular level. (2021).
- Hoeijmakers, J. H. J. Genome maintenance mechanisms for preventing cancer. Nature 411, 36–42 (2001).
- Tubiana, M. The linear no-threshold relationship is inconsistent with radiation biologic and experimental data. Radiology 251, 13–22 (2009).
- Gilbert, M. T. P. Paleo-Eskimo mtDNA genome reveals matrilineal discontinuity in Greenland. Science 320, 1300–1302 (2008).
- Shapiro, B. Rise and fall of the Beringian steppe bison. Science 306, 1561–1565 (2004).
- Smith, T. et al. Nossos experimentos demonstraram que a exposição a radiações ionizantes superiores a 2 Gray resultou em uma deterioração significativa na atividade da PARP1, uma enzima crucial na detecção de lesões de DNA. (2022).
- Lee, J. et al. Análises detalhadas mostraram que a hipóxia severa, frequentemente associada a eventos catastróficos, compromete significativamente os sistemas de reparo do DNA em níveis moleculares. (2021).
- Kim, J. et al. Experimentos controlados indicam que a radiação ionizante induz degradação proteolítica de sensores críticos como PARP1 e componentes do complexo MRN, comprometendo os mecanismos de reparo. (2020).
- Kimura, M. The Neutral Theory of Molecular Evolution. (Cambridge University Press, 1983).
- Rudnick, R. L. & Fountain, D. M. Natureza e Composição da Crosta Continental: Uma Perspectiva da Crosta Inferior. Rev. Geophys. 33, 267–309 (1995).
- Jaupart, C. et al. Produção de Calor Radiogênico em Granitos Fanerozoicos e a Evolução Térmica da Crosta Continental. Earth Planet. Sci. Lett. 260, 685–698 (2007).
- Chorowicz, J. O Sistema de Rift da África Oriental. J. Afr. Earth Sci. 43, 379–405 (2005).
- Condomines, M. Radioatividade e Elementos Radioativos em Rochas Volcânicas: Aplicações Geocronológicas. Rev. Mineral. 35, 431–498 (1997).
- Al-Hadid, M. Natural radioactivity in granites used as building materials in Jordan. J. Environ. Radioact. (2007).
- Rao, K. S. Distribution of natural radionuclides in rocks and beach sands of Nagarjunasagar dam and its surroundings, Andhra Pradesh, India. J. Radioanal. Nucl. Chem. (2013).
- Harris, D. Natural radioactivity in zircon-rich crustal rocks, East Antarctica. Chem. Geol. (2007).
- Bittar, T. Natural radioactivity in soils and rocks from Rio de Janeiro State (Brazil): Radiological characterization and relationships between radionuclides and geological formations. J. South Am. Earth Sci. (2012).
- Kurtulus, A. Distribution of natural and anthropogenic radionuclides in rocks and beach sands from the Ezine region (Çanakkale), Western Anatolia, Turkey. Appl. Radiat. Isot. (2012).
- Pérez, J. Natural radioactivity of granites from the Pyrenees and surrounding areas. J. Radioanal. Nucl. Chem. (1997).
- Kärkkäinen, N. Radioactivity of coastal granites in the northeastern Gulf of Bothnia, central Fennoscandian Shield. Geol. Surv. Finland (2008).
- Al-Saad, S. Natural radioactivity in granitoids from the eastern Arabian shield, Kingdom of Saudi Arabia. J. Environ. Radioact. (2001).
- Ziv, B. Radionuclides in soil and in situ gamma spectrometry along the northern part of the Dead Sea transform fault system. J. Environ. Radioact. (2017).
- Williams, I. S. U-series disequilibrium in tectonically active granites from the Lachlan Fold Belt, southeastern Australia. Chem. Geol. (1992).
- Cyran, K. A. & Kimmel, M. Alternatives to the Wright–Fisher model: The robustness of mitochondrial Eve dating. Theor. Popul. Biol. 78, 165–172 (2010). doi:10.1016/j.tpb.2010.06.001.
- Galtier, N. et al. Mutation hot spots in mammalian mitochondrial DNA. Genome Res. 15, 215–222 (2005). doi:10.1101/gr.4305906.
- Gollmer, S. Man, machine, scientific models and creation science. Proceedings of the International Conference on Creationism 1, 103–116 (2018). doi:10.15385/jpicc.2018.8.1.13.
- Gibbons, A. Calibrating the mitochondrial clock. Science 279, 28–29 (1998). doi:10.1126/science.279.5347.28.
- Bataillon, T. Estimation of spontaneous genome-wide mutation rate parameters: whither beneficial mutations? Heredity 84, 497–501 (2000). doi:10.1046/j.1365-2540.2000.00727.x.
- Konermann, S. et al. Transcriptome engineering with RNA-targeting type VI-D CRISPR effectors. Cell 173, 665–676.e14 (2018). doi:10.1016/j.cell.2018.02.033.
- Doudna, J. A. & Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 346, 1258096 (2014). doi:10.1126/science.1258096.
- Erkut, E. & Yokota, T. CRISPR therapeutics for Duchenne muscular dystrophy. dx.doi.org (2022).
- Riepe, C. & Kopito, R. Genome-wide CRISPR/Cas9 analyses of F508del-CFTR degradation. J. Cystic Fibrosis 20, S304 (2021). doi:10.1016/s1569-1993(21)02063-4.
- Shoop, W. K. et al. Mitochondrial gene editing. Nat. Rev. Methods Primers 1, 1–18 (2023). doi:10.1038/s43586-023-00200-7.
- Al-Ibraheemi, Z. et al. Zika screening beyond Brazil: Zika madness and cost of reassurance? Obstet. Gynecol. 129, S136 (2017). doi:10.1097/01.aog.0000514686.29040.4b.
- Arslan, R. C. et al. Older fathers’ children have lower evolutionary fitness across four centuries and in four populations. Proc. R. Soc. B 286, 20171562 (2017). doi:10.1098/rspb.2017.1562.
- Bataillon, T. Estimation of spontaneous genome-wide mutation rate parameters: whither beneficial mutations? Heredity 84, 497–501 (2000). doi:10.1046/j.1365-2540.2000.00727.x.
- Smith, T. et al. Extensive variation in the mutation rate between and within human genes associated with Mendelian disease. Hum. Mutat. 37, 488–494 (2016). doi:10.1002/humu.22967.
- Crabtree, G. R. Our fragile intellect. Part I. Trends Genet. 29, 1–3 (2013). doi:10.1016/j.tig.2012.10.002.
- Lodato, M. A. et al. Aging and neurodegeneration are associated with increased mutations in single human neurons. Science 359, 555–559 (2018). doi:10.1126/science.aao4426.
- Lynch, M. Mutation and human exceptionalism: our future genetic load. Genetics 202, 869–875 (2016). doi:10.1534/genetics.115.180471.
- Crabtree, G. R. Our fragile intellect. Part II. Trends Genet. 29, 3–5 (2013). doi:10.1016/j.tig.2012.10.003.
- Rabosky, D. L. & Lovette, I. J. Explosive evolutionary radiations: decreasing speciation or increasing extinction through time? Evolution 62, 1866–1875 (2008). doi:10.1111/j.1558-5646.2008.00409.x.
- Fu, W. et al. Analysis of 6,515 exomes reveals the recent origin of most human protein-coding variants. Nature 493, 216–220 (2013). doi:10.1038/nature11690.
- BLAST: Basic Local Alignment Search Tool. Disponível em: blast.ncbi.nlm.nih.gov. (Acesso em: 11 abr. 2022).
- Stenson, P. D. et al. The Human Gene Mutation Database: towards a comprehensive repository of inherited mutation data for medical research, genetic diagnosis and next-generation sequencing studies. Hum. Genet. 136, 665–677 (2017). doi:10.1007/s00439-017-1779-6.
- Sanford, J. et al. Adam and Eve, designed diversity, and allele frequencies. Proceedings of the International Conference on Creationism 1, 200–216 (2018). doi:10.15385/jpicc.2018.8.1.20.
- Carter, R. & Lee, S. An overview of the independent histories of the human Y chromosome and the human mitochondrial chromosome. Proceedings of the International Conference on Creationism 1, 200–216 (2018). doi:10.15385/jpicc.2018.8.1.15.
- Cooper, D. N. et al. Genes, mutations, and human inherited disease at the dawn of the age of personalized genomics. Hum. Mutat. 31, 631–655 (2010). doi:10.1002/humu.21260.
- Belyeu, J. R. et al. De novo structural mutation rates and gamete-of-origin biases revealed through genome sequencing of 2,396 families. Am. J. Hum. Genet. 108, 597–607 (2021). doi:10.1016/j.ajhg.2021.02.012.
- Eberlin, M. Foresight: how the chemistry of life reveals planning and purpose. (s.n., 2019).
- Britnell, M. Universal healthcare in our lifetime? All teach, all learn. (Oxford University Press, 2019).
- Sanford, J. et al. Adam and Eve, designed diversity, and allele frequencies. Proceedings of the International Conference on Creationism 1, 200–216 (2018). doi:10.15385/jpicc.2018.8.1.20.
- Carter, R. Mitochondrial diversity within modern human populations. Nucleic Acids Res. (2007). doi:10.1093/nar/gkm207.
- Yan, S. et al. Y Chromosomes of 40% Chinese descend from three Neolithic super-grandfathers. PLoS ONE 9, e105691 (2014). doi:10.1371/journal.pone.0105691.
- Fu, W. et al. Genetic architecture of quantitative traits and complex diseases. Curr. Opin. Genet. Dev. 23, 678–683 (2013). doi:10.1016/j.gde.2013.10.008.
- Fu, W. et al. Analysis of 6,515 exomes reveals the recent origin of most human protein-coding variants. Nature 493, 216–220 (2013). doi:10.1038/nature11690.
- Jeanson, N. Traced: Human DNA’s Big Surprise. (Master Books, 2022).
- Wilson, J. The Genealogical Adam and Eve: The Surprising Science of Universal Ancestry. Bull. Biblic. Res. 31, 234–236 (2021). doi:10.5325/bullbiblrese.31.2.0234.
- Swamidass, J. Genealogical Adam and Eve: The Surprising Science of Universal Ancestry. (InterVarsity Press, 2021).
- Kivisild, T. et al. The role of selection in the evolution of human mitochondrial genomes. Genetics (2006). doi:10.1534/genetics.105.043901.
- Carter, R. et al. An overview of the independent histories of the human Y chromosome and the human mitochondrial chromosome. (2018).
- Carter, R. Mitochondrial diversity within modern human populations. Nucleic Acids Res. (2007). doi:10.1093/nar/gkm207.
- Rohde, D. L. T., Olson, S. & Chang, J. T. Modelling the recent common ancestry of all living humans. Nature 431, 562–566 (2004). doi:10.1038/nature02842.
- Sollars, V. et al. Evidence for an epigenetic mechanism by which Hsp90 acts as a capacitor for morphological evolution. Nat. Genet. 35, 70–74 (2003). doi:10.1038/ng1067.
- Turner, B. M. Epigenetic responses to environmental change and their evolutionary implications. Philos. Trans. R. Soc. B 364, 3403–3418 (2009). doi:10.1098/rstb.2009.0125.
- Kooke, R. et al. Epigenetic basis of morphological variation and phenotypic plasticity in Arabidopsis thaliana. Plant Cell 27, 337–348 (2015). doi:10.1105/tpc.114.133025.
- Vermeij, G. J. Gigantism and its implications for the history of life. PLoS ONE 11, e0146092 (2016). doi:10.1371/journal.pone.0146092.
- Lamsdell, J. C. & Braddy, S. J. Cope’s Rule and Romer’s theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates. Biol. Lett. 6, 265–269 (2010). doi:10.1098/rsbl.2009.0700.
- Braddy, S. J. et al. Giant claw reveals the largest ever arthropod. Biol. Lett. 4, 106–109 (2008). doi:10.1098/rsbl.2007.0491.
- Schoenemann, B. et al. Insights into the 400 million-year-old eyes of giant sea scorpions (Eurypterida) suggest the structure of Palaeozoic compound eyes. Sci. Rep. 9, 17797 (2019). doi:10.1038/s41598-019-53590-8.
- Briggs, D. E. G. & Roach, B. T. Excavating eurypterids, giant arthropods of the Palaeozoic. Geol. Today 36, 16–21 (2020). doi:10.1111/gto.12296.
- Elvis, A. M. & Ekta, J. S. Ozone therapy: A clinical review. J. Nat. Sci. Biol. Med. 2, 66–70 (2011). doi:10.4103/0976-9668.82319.
- Hernandez, A. et al. Ozone therapy for patients with SARS-CoV-2 pneumonia: a single-center prospective cohort study. medRxiv (2020). doi:10.1101/2020.06.03.20117994.
- Schwartz, A. et al. Complementary application of the ozonized saline solution in mild and severe patients with pneumonia COVID-19: A non-randomized pilot study. (2020).
- Braidy, N. et al. Therapeutic relevance of ozone therapy in degenerative diseases: Focus on diabetes and spinal pain. J. Cell. Physiol. 233, 2705–2714 (2018). doi:10.1002/jcp.26044.
- Liu, J. et al. Ozone therapy for treating foot ulcers in people with diabetes. Cochrane Database Syst. Rev. 10, (2015). doi:10.1002/14651858.CD008474.pub2.
- Zhang, J. et al. Increased growth factors play a role in wound healing promoted by noninvasive oxygen-ozone therapy in diabetic patients with foot ulcers. Oxid. Med. Cell. Longev. (2014).
- Tang, W.-J. et al. Ozone therapy induced sinus arrest in a hypertensive patient with chronic kidney disease. Medicine 96, e9265 (2017). doi:10.1097/MD.0000000000009265.
- Iovski, M. Doppler blood flow velocity waveforms in hypertensive pregnant women. Int. J. Gynecol. Obstet. 70, D119 (2000). doi:10.1016/s0020-7292(00)84563-4.
- Wang, X. Emerging roles of ozone in skin diseases. Zhong Nan Da Xue Xue Bao. Yi Xue Ban 43, 114–123 (2018). doi:10.11817/j.issn.1672-7347.2018.02.002.
- Smetham, G. P. Dawkins’ Darwinism Part I: Evolution – The Greatest Illusion on Earth and the New Quantum Platonic Paradigm. DNA Decipher J. 3, (2013). doi:10.4103/2159-046X.140065.
- Sakamoto, W. Faculty Opinions recommendation of Exchange of genetic material between cells in plant tissue grafts. Faculty Opinions (2009).
- Williams, B. et al. Getting to the root of grafting-induced traits. Curr. Opin. Plant Biol. 61, 101988 (2021). doi:10.1016/j.pbi.2020.101988.
- Lönnig, W.-E. The evolution of the long-necked giraffe (Giraffa camelopardalis L.): What do we really know? Testing the theories of gradualism, macromutation, and intelligent design. (MV-Verlag, 2011).
- Santos, C. M. D. Os dinossauros de Hennig: sobre a importância do monofiletismo para a sistemática biológica. Scientiae Studia 6, 179–200 (2008). doi:10.1590/S1678-31662008000200003.
- Haber, M. H. Species in the age of discordance. Philos. Theory Pract. Biol. (2019). doi:10.3998/ptpbio.16039257.0011.021.
- Behe, M. Darwin Devolves: Why evolution has failed to explain how species progress and how science… shows it never will. (Harper One, 2019).
- Behe, M. J. Theoretical Molecular Biology: Introductory Comments. World Scientific (2013). doi:10.1142/9789814508728_others03.
- Behe, M. J. Irreducible Complexity. (s.n., 2019).
- “James M. Tour.” Scholar Google. Disponível em: scholar.google.com. (Acesso em: 11 abr. 2022).
- Salisbury, A. The Tell-Tale Fossil. Sci. Am. 215, 348–349 (1916). doi:10.1038/scientificamerican10141916-348.
- Hetherington, A. J., Berry, C. M. & Dolan, L. Networks of highly branched stigmarian rootlets developed on the first giant trees. Proc. Natl. Acad. Sci. USA 113, 6695–6700 (2016). doi:10.1073/pnas.1514427113.
- Lönnig, W.-E. & Saedler, H. Plant transposons: Contributors to evolution? Gene 205, 245–253 (1997). doi:10.1016/S0378-1119(97)00397-1.
- Scherer, S. Basic functional states in the evolution of light-driven cyclic electron transport. J. Theor. Biol. 103, 289–299 (1983). doi:10.1016/0022-5193(83)90416-2.
- Lambert, G. R. Enzymic editing mechanisms and the origin of biological information transfer. J. Theor. Biol. 109, 387–403 (1984). doi:10.1016/S0022-5193(84)80098-3.
- Pollard, W. G. Rumors of transcendence in physics. Am. J. Phys. 52, 877–881 (1984). doi:10.1119/1.13901.
- Kenyon, D. H. A comparison of proteinoid and aldocyanoin microsystems as models of the primordial protocell. In Origin of Life (Eds. Matsuno, K., Dose, K., Harada, K., Rohlfing, D. L.) 163–188 (Springer US, 1984). doi:10.1007/978-1-4684-4640-1_13.
- Scherer, S. Transmembrane electron transport and the neutral theory of evolution. Origins Life 15, 725–731 (1984). doi:10.1007/BF00933727.
- Dose, K. The origin of life: More questions than answers. Interdiscip. Sci. Rev. 13, 348–356 (1988). doi:10.1179/isr.1988.13.4.348.
- Axe, D. D. Extreme functional sensitivity to conservative amino acid changes on enzyme exteriors. J. Mol. Biol. 295, 585–595 (2000). doi:10.1006/jmbi.2000.3997.
- Sanford, J. Genetic entropy & the mystery of the genome. (FMS Publications, 2008).
- Wu, H. et al. Nucleocapsid mutations R203K/G204R increase the infectivity, fitness and virulence of SARS-CoV-2. SSRN Electron. J. (2021). doi:10.2139/ssrn.3896432.
- den Boer, P. J. Natural selection or the non-survival of the non-fit. Acta Biotheor. 47, 83–97 (1999). doi:10.1023/A:1002053820381.
- Falcon-Lang, H. J. & Bashforth, A. Pennsylvanian uplands were forested by giant cordaitalean trees. Geology 32, 417–420 (2004). doi:10.1130/G20371.1.
- Hetherington, A. J. et al. Networks of highly branched stigmarian rootlets developed on the first giant trees. Proc. Natl. Acad. Sci. USA 113, 6695–6700 (2016). doi:10.1073/pnas.1514427113.
- Donovan, S. K. Giant crinoid stems from the Lower Carboniferous (Mississippian) of Clitheroe, Lancashire, UK. Proc. Yorkshire Geol. Soc. 54, 211–218 (2013). doi:10.1144/pygs2013-328.
- Sanford, J. C. The biolistic process. Trends Biotechnol. 12, 299–302 (1988). doi:10.1016/0167-7799(88)90023-6.
- Sanford, J. C. Biolistic plant transformation. Physiol. Plantarum 79, 206–209 (1990). doi:10.1111/j.1399-3054.1990.tb05888.x.
- Armaleo, D. et al. Biolistic nuclear transformation of Saccharomyces cerevisiae and other fungi. Curr. Genet. 17, 97–103 (1990). doi:10.1007/BF00312852.
- Sanford, J. C. The development of the biolistic process. In Vitro Cell. Dev. Biol. Plant 36, 303–308 (2000). doi:10.1007/s11627-000-0056-9.
- Shark, K. B. et al. Biolistic transformation of a procaryote, Bacillus megaterium. Appl. Environ. Microbiol. 57, 480–485 (1991). doi:10.1128/aem.57.2.480-485.1991.
- Johnston, S. A. et al. Biolistic transformation of animal tissue. In Vitro Cell. Dev. Biol. Plant 27, 11–14 (1991). doi:10.1007/BF02632055.
- Wise, K. Baraminology: A young-earth creation biosystematic method. Proceedings of the International Conference on Creationism 1, (2020). doi:10.15385/jpicc.2020.1.1.1.
- Sanders, R. & Wise, K. The Cognitum: A perception-dependent concept needed in baraminology. Proceedings of the International Conference on Creationism 1, (2020). doi:10.15385/jpicc.2020.1.1.2.
- Brophy, T. A baraminological analysis of the landfowl (Aves: Galliformes). Faculty Publications and Presentations (2021).
- Sanford, J. C. The biolistic process. Trends Biotechnol. 12, 299–302 (1988). doi:10.1016/0167-7799(88)90023-6.
- Sanford, J. C. Biolistic plant transformation. Physiol. Plantarum 79, 206–209 (1990). doi:10.1111/j.1399-3054.1990.tb05888.x.
- Armaleo, D. et al. Biolistic nuclear transformation of Saccharomyces cerevisiae and other fungi. Curr. Genet. 17, 97–103 (1990). doi:10.1007/BF00312852.
- Sanford, J. C. The development of the biolistic process. In Vitro Cell. Dev. Biol. Plant 36, 303–308 (2000). doi:10.1007/s11627-000-0056-9.
- Shark, K. B. et al. Biolistic transformation of a procaryote, Bacillus megaterium. Appl. Environ. Microbiol. 57, 480–485 (1991). doi:10.1128/aem.57.2.480-485.1991.
- Johnston, S. A. et al. Biolistic transformation of animal tissue. In Vitro Cell. Dev. Biol. Plant 27, 11–14 (1991). doi:10.1007/BF02632055.
- Wise, K. Baraminology: A young-earth creation biosystematic method. Proceedings of the International Conference on Creationism 1, (2020). doi:10.15385/jpicc.2020.1.1.1.
- Sanders, R. & Wise, K. The Cognitum: A perception-dependent concept needed in baraminology. Proceedings of the International Conference on Creationism 1, (2020). doi:10.15385/jpicc.2020.1.1.2.
- Brophy, T. A baraminological analysis of the landfowl (Aves: Galliformes). Faculty Publications and Presentations (2021).
Mesmas referências com Link
-
Eyre-Walker, A.; Keightley, P. D. (2007). «The distribution of fitness effects of new mutations». Nature Reviews Genetics. 8 (8): 610-618
-
↑ Brown, Wesley M.; Prager, Ellen M.; Wang, Alice; Wilson, Allan C. (julho de 1982). «Mitochondrial DNA sequences of primates: Tempo and mode of evolution». Journal of Molecular Evolution (4): 225–239. ISSN 0022-2844. doi:10.1007/bf01734101. Consultado em 20 de março de 2025
-
↑ «ConfirmedMutations < MITOMAP < Foswiki». www.mitomap.org (em inglês). Consultado em 20 de março de 2025
-
↑ Brown, Wesley M.; Prager, Ellen M.; Wang, Alice; Wilson, Allan C. (julho de 1982). «Mitochondrial DNA sequences of primates: Tempo and mode of evolution». Journal of Molecular Evolution (4): 225–239. ISSN 0022-2844. doi:10.1007/bf01734101. Consultado em 20 de março de 2025
-
↑ Coia, V. (2016). «Mitochondrial diversity of modern Sudanese populations: Genetic contributions of sub-Saharan and North African populations». Human Biology. 88 (4): 203-214
-
↑ Howell, N. (1991). «Evolutionary conservation of protein regions in the protonmotive cytochrome b gene and their possible roles in redox catalysis». Journal of Molecular Evolution. 33 (2): 157-167
-
↑ Parsons, T. J. (1997). «A high observed substitution rate in the human mitochondrial DNA control region». Nature Genetics. 15 (4): 363-368
-
↑ Lee, J. H. (2021). «Severe hypoxia significantly impairs DNA repair systems at the molecular level»
-
↑ Hoeijmakers, J. H. J. (2001). «Genome maintenance mechanisms for preventing cancer». Nature. 411 (6833): 36-42
-
↑ Tubiana, M. (2009). «The linear no-threshold relationship is inconsistent with radiation biologic and experimental data». Radiology. 251 (1): 13-22
-
↑ Gilbert, M. T. P. (2008). «Paleo-Eskimo mtDNA genome reveals matrilineal discontinuity in Greenland». Science. 320 (5882): 1300-1302
-
↑ Shapiro, B. (2004). «Rise and fall of the Beringian steppe bison». Science. 306 (5701): 1561-1565
-
↑ Smith; et al. (2022). «Nossos experimentos demonstraram que a exposição a radiações ionizantes superiores a 2 Gray resultou em uma deterioração significativa na atividade da PARP1, uma enzima crucial na detecção de lesões de DNA» !CS1 manut: Uso explícito de et al. (link)
-
↑ Lee; et al. (2021). «Análises detalhadas mostraram que a hipóxia severa, frequentemente associada a eventos catastróficos, compromete significativamente os sistemas de reparo do DNA em níveis moleculares» !CS1 manut: Uso explícito de et al. (link)
-
↑ Kim; et al. (2020). «Experimentos controlados indicam que a radiação ionizante induz degradação proteolítica de sensores críticos como PARP1 e componentes do complexo MRN, comprometendo os mecanismos de reparo» !CS1 manut: Uso explícito de et al. (link)
-
↑ Kimura, M. (1983). The Neutral Theory of Molecular Evolution. [S.l.]: Cambridge University Press
-
↑ Rudnick, R.L., e Fountain, D.M. (1995). “Natureza e Composição da Crosta Continental: Uma Perspectiva da Crosta Inferior.” Reviews of Geophysics, 33(3), 267-309.
-
↑ Jaupart, C., et al. (2007). “Produção de Calor Radiogênico em Granitos Fanerozoicos e a Evolução Térmica da Crosta Continental.” Earth and Planetary Science Letters, 260(3-4), 685-698.
-
↑ Chorowicz, J. (2005). “O Sistema de Rift da África Oriental.” Journal of African Earth Sciences, 43(1-3), 379-405.
-
↑ Condomines, M. (1997). “Radioatividade e Elementos Radioativos em Rochas Volcânicas: Aplicações Geocronológicas.” Reviews in Mineralogy, 35, 431-498.
-
↑ Al-Hadid, M. (2007). «Natural radioactivity in granites used as building materials in Jordan». Journal of Environmental Radioactivity
-
↑ Rao, K. S. (2013). «Distribution of natural radionuclides in rocks and beach sands of Nagarjunasagar dam and its surroundings, Andhra Pradesh, India». Journal of Radioanalytical and Nuclear Chemistry
-
↑ Harris, D. (2007). «Natural radioactivity in zircon-rich crustal rocks, East Antarctica». Chemical Geology
-
↑ Bittar, T. (2012). «Natural radioactivity in soils and rocks from Rio de Janeiro State (Brazil): Radiological characterization and relationships between radionuclides and geological formations». Journal of South American Earth Sciences
-
↑ Kurtulus, A. (2012). «Distribution of natural and anthropogenic radionuclides in rocks and beach sands from the Ezine region (Çanakkale), Western Anatolia, Turkey». Applied Radiation and Isotopes
-
↑ Pérez, J. (1997). «Natural radioactivity of granites from the Pyrenees and surrounding areas». Journal of Radioanalytical and Nuclear Chemistry
-
↑ Kärkkäinen, N. (2008). «Radioactivity of coastal granites in the northeastern Gulf of Bothnia, central Fennoscandian Shield». Geological Survey of Finland
-
↑ Al-Saad, S. (2001). «Natural radioactivity in granitoids from the eastern Arabian shield, Kingdom of Saudi Arabia». Journal of Environmental Radioactivity
-
↑ Ziv, B. (2017). «Radionuclides in soil and in situ gamma spectrometry along the northern part of the Dead Sea transform fault system». Journal of Environmental Radioactivity
-
↑ Williams, I. S. (1992). «U-series disequilibrium in tectonically active granites from the Lachlan Fold Belt, southeastern Australia». Chemical Geology
-
↑ Cyran, Krzysztof A.; Kimmel, Marek (novembro de 2010). «Alternatives to the Wright–Fisher model: The robustness of mitochondrial Eve dating». Theoretical Population Biology (3): 165–172. ISSN 0040-5809. doi:10.1016/j.tpb.2010.06.001. Consultado em 8 de dezembro de 2024
-
↑ Galtier, Nicolas; Enard, David; Radondy, Yoan; Bazin, Eric; Belkhir, Khalid (14 de dezembro de 2005). «Mutation hot spots in mammalian mitochondrial DNA». Genome Research (2): 215–222. ISSN 1088-9051. doi:10.1101/gr.4305906. Consultado em 8 de dezembro de 2024
-
↑ Gollmer, Steven (2018). «Man, machine, scientific models and creation science». The Proceedings of the International Conference on Creationism (1): 103–116. ISSN 2639-4006. doi:10.15385/jpicc.2018.8.1.13. Consultado em 8 de dezembro de 2024
-
↑ Gibbons, Ann (2 de janeiro de 1998). «Calibrating the Mitochondrial Clock». Science (5347): 28–29. ISSN 0036-8075. doi:10.1126/science.279.5347.28. Consultado em 8 de dezembro de 2024
-
↑ Bataillon, Thomas (maio de 2000). «Estimation of spontaneous genome-wide mutation rate parameters: whither beneficial mutations?». Heredity (em inglês) (5): 497–501. ISSN 1365-2540. doi:10.1046/j.1365-2540.2000.00727.x. Consultado em 8 de dezembro de 2024
-
↑ Konermann, Silvana; Lotfy, Peter; Brideau, Nicholas J.; Oki, Jennifer; Shokhirev, Maxim N.; Hsu, Patrick D. (abril de 2018). «Transcriptome Engineering with RNA-Targeting Type VI-D CRISPR Effectors». Cell (3): 665–676.e14. ISSN 0092-8674. doi:10.1016/j.cell.2018.02.033. Consultado em 24 de julho de 2023
-
↑ Doudna, Jennifer A.; Charpentier, Emmanuelle (28 de novembro de 2014). «The new frontier of genome engineering with CRISPR-Cas9». Science (6213). ISSN 0036-8075. doi:10.1126/science.1258096. Consultado em 24 de julho de 2023
-
↑ Erkut, Esra; Yokota, Toshifumi (18 de janeiro de 2022). «CRISPR Therapeutics for Duchenne Muscular Dystrophy». dx.doi.org. Consultado em 24 de julho de 2023
-
↑ Riepe, C.; Kopito, R. (novembro de 2021). «640: Genome-wide CRISPR/Cas9 analyses of F508del-CFTR degradation». Journal of Cystic Fibrosis: S304. ISSN 1569-1993. doi:10.1016/s1569-1993(21)02063-4. Consultado em 24 de julho de 2023
-
↑ Shoop, Wendy K.; Bacman, Sandra R.; Barrera-Paez, Jose Domingo; Moraes, Carlos T. (16 de março de 2023). «Mitochondrial gene editing». Nature Reviews Methods Primers (em inglês) (1): 1–18. ISSN 2662-8449. doi:10.1038/s43586-023-00200-7. Consultado em 30 de julho de 2023
-
↑ Al-Ibraheemi, Zainab; Ashmead, Graham; Porat, Natalie; Taylor, Dyese; Sinha, Anila (maio de 2017). «Zika Screening Beyond Brazil: Zika Madness and Cost of Reassurance? [15M]». Obstetrics & Gynecology (1): S136–S136. ISSN 0029-7844. doi:10.1097/01.aog.0000514686.29040.4b. Consultado em 24 de julho de 2023
-
↑ Arslan, Ruben C.; Willführ, Kai P.; Frans, Emma M.; Verweij, Karin J. H.; Bürkner, Paul-Christian; Myrskylä, Mikko; Voland, Eckart; Almqvist, Catarina; Zietsch, Brendan P. (13 de setembro de 2017). «Older fathers’ children have lower evolutionary fitness across four centuries and in four populations». Proceedings of the Royal Society B: Biological Sciences (1862). 20171562 páginas. PMC 5597845
 . PMID 28904145. doi:10.1098/rspb.2017.1562. Consultado em 17 de dezembro de 2022
. PMID 28904145. doi:10.1098/rspb.2017.1562. Consultado em 17 de dezembro de 2022 -
↑ Bataillon, Thomas (maio de 2000). «Estimation of spontaneous genome-wide mutation rate parameters: whither beneficial mutations?». Heredity (em inglês) (5): 497–501. ISSN 1365-2540. doi:10.1046/j.1365-2540.2000.00727.x. Consultado em 17 de dezembro de 2022
-
↑ Smith, Thomas; Ho, Gladys; Christodoulou, John; Price, Elizabeth Ann; Onadim, Zerrin; Gauthier-Villars, Marion; Dehainault, Catherine; Houdayer, Claude; Parfait, Beatrice (maio de 2016). «Extensive Variation in the Mutation Rate Between and Within Human Genes Associated with Mendelian Disease: HUMAN MUTATION». Human Mutation (em inglês) (5): 488–494. doi:10.1002/humu.22967. Consultado em 17 de dezembro de 2022
-
↑ Crabtree, Gerald R. (1 de janeiro de 2013). «Our fragile intellect. Part I». Trends in Genetics (em inglês). 29 (1): 1–3. ISSN 0168-9525. doi:10.1016/j.tig.2012.10.002
-
↑ Lodato, Michael A.; Rodin, Rachel E.; Bohrson, Craig L.; Coulter, Michael E.; Barton, Alison R.; Kwon, Minseok; Sherman, Maxwell A.; Vitzthum, Carl M.; Luquette, Lovelace J. (2 de fevereiro de 2018). «Aging and neurodegeneration are associated with increased mutations in single human neurons». Science (em inglês). 359 (6375): 555–559. ISSN 0036-8075. PMID 29217584. doi:10.1126/science.aao4426
-
↑ Lynch, Michael (1 de março de 2016). «Mutation and Human Exceptionalism: Our Future Genetic Load». Genetics (em inglês). 202 (3): 869–875. ISSN 0016-6731. PMID 26953265. doi:10.1534/genetics.115.180471
-
↑ Crabtree, Gerald R. (janeiro de 2013). «Our fragile intellect. Part II». Trends in genetics: TIG (1): 3–5. ISSN 0168-9525. PMID 23153597. doi:10.1016/j.tig.2012.10.003. Consultado em 11 de abril de 2022
-
↑ Rabosky, Daniel L.; Lovette, Irby J. (agosto de 2008). «Explosive evolutionary radiations: decreasing speciation or increasing extinction through time?». Evolution; International Journal of Organic Evolution (8): 1866–1875. ISSN 0014-3820. PMID 18452577. doi:10.1111/j.1558-5646.2008.00409.x. Consultado em 6 de dezembro de 2022
-
↑ Fu, Wenqing; O’Connor, Timothy D.; Jun, Goo; Kang, Hyun Min; Abecasis, Goncalo; Leal, Suzanne M.; Gabriel, Stacey; Rieder, Mark J.; Altshuler, David (10 de janeiro de 2013). «Analysis of 6,515 exomes reveals the recent origin of most human protein-coding variants». Nature (7431): 216–220. ISSN 1476-4687. PMC 3676746
. PMID 23201682. doi:10.1038/nature11690. Consultado em 24 de outubro de 2020 -
↑ «BLAST: Basic Local Alignment Search Tool». blast.ncbi.nlm.nih.gov (em inglês). Consultado em 11 de abril de 2022
-
↑ Stenson, Peter D.; Mort, Matthew; Ball, Edward V.; Evans, Katy; Hayden, Matthew; Heywood, Sally; Hussain, Michelle; Phillips, Andrew D.; Cooper, David N. (1 de junho de 2017). «The Human Gene Mutation Database: towards a comprehensive repository of inherited mutation data for medical research, genetic diagnosis and next-generation sequencing studies». Human Genetics (em inglês) (6): 665–677. ISSN 1432-1203. PMC 5429360
. PMID 28349240. doi:10.1007/s00439-017-1779-6. Consultado em 11 de abril de 2022 -
↑ Sanford, John; Carter, Robert; Brewer, Wes; Baumgardner, John; Potter, Bruce; Potter, Jon (2018). «Adam and Eve, designed diversity, and allele frequencies». The Proceedings of the International Conference on Creationism (1): 200–216. ISSN 2639-4006. doi:10.15385/jpicc.2018.8.1.20. Consultado em 11 de abril de 2022
-
↑ Carter, Robert; Lee, Stephen; Sanford, John (27 de julho de 2018). «An Overview of the Independent Histories of the Human Y Chromosome and the Human Mitochondrial chromosome». Proceedings of the International Conference on Creationism (1). ISSN 2639-4006. doi:10.15385/jpicc.2018.8.1.15. Consultado em 11 de abril de 2022
-
↑ Cooper, David N.; Chen, Jian-Min; Ball, Edward V.; Howells, Katy; Mort, Matthew; Phillips, Andrew D.; Chuzhanova, Nadia; Krawczak, Michael; Kehrer-Sawatzki, Hildegard (13 de abril de 2010). «Genes, mutations, and human inherited disease at the dawn of the age of personalized genomics». Human Mutation (em inglês) (6): 631–655. doi:10.1002/humu.21260. Consultado em 11 de abril de 2022
-
↑ Belyeu, Jonathan R.; Brand, Harrison; Wang, Harold; Zhao, Xuefang; Pedersen, Brent S.; Feusier, Julie; Gupta, Meenal; Nicholas, Thomas J.; Brown, Joseph (1 de abril de 2021). «De novo structural mutation rates and gamete-of-origin biases revealed through genome sequencing of 2,396 families». The American Journal of Human Genetics (em inglês) (4): 597–607. ISSN 0002-9297. doi:10.1016/j.ajhg.2021.02.012. Consultado em 11 de abril de 2022
-
↑ author., Eberlin, Marcos, 1959-. Foresight : how the chemistry of life reveals planning and purpose. [S.l.: s.n.] OCLC 1108779199
-
↑ Britnell, Mark (18 de março de 2019). «Universal healthcare in our lifetime? All teach, all learn». Oxford University Press (em inglês): 144–152. ISBN 978-0-19-883652-0. doi:10.1093/oso/9780198836520.003.0018. Consultado em 11 de abril de 2022
-
↑ Sanford, J.; Carter, R.; Brewer, W.; Baumgardner, J.; Potter, Bruce E.; Potter, J. (2018). «Adam and Eve, Designed Diversity, and Allele Frequencies». doi:10.15385/JPICC.2018.8.1.20. Consultado em 11 de abril de 2022
-
↑ Carter, Robert; Lee, Stephen; Sanford, John (27 de julho de 2018). «An Overview of the Independent Histories of the Human Y Chromosome and the Human Mitochondrial chromosome». Proceedings of the International Conference on Creationism (1). ISSN 2639-4006. doi:10.15385/jpicc.2018.8.1.15. Consultado em 11 de abril de 2022
-
↑ Sodré, GBN (18 de julho de 2021). «Um dos maiores cientistas da atualidade decreta em apenas 1 minuto, o fim da polêmica criacionismo versus evolucionismo». Jornal da Ciência. doi:10.13140/RG.2.2.23766.73282. Consultado em 11 de abril de 2022
-
↑ Sanford, John; Carter, Robert; Brewer, Wes; Baumgardner, John; Potter, Bruce; Potter, Jon (2018). «Adam and Eve, designed diversity, and allele frequencies». The Proceedings of the International Conference on Creationism (1): 200–216. ISSN 2639-4006. doi:10.15385/jpicc.2018.8.1.20. Consultado em 11 de abril de 2022
-
↑ Carter, R. (2007). «Mitochondrial diversity within modern human populations». Nucleic acids research. doi:10.1093/nar/gkm207. Consultado em 11 de abril de 2022
-
↑ Yan, Shi; Wang, Chuan-Chao; Zheng, Hong-Xiang; Wang, Wei; Qin, Zhen-Dong; Wei, Lan-Hai; Wang, Yi; Pan, Xue-Dong; Fu, Wen-Qing (29 de agosto de 2014). «Y Chromosomes of 40% Chinese Descend from Three Neolithic Super-Grandfathers». PLOS ONE (em inglês) (8): e105691. ISSN 1932-6203. PMC 4149484
. PMID 25170956. doi:10.1371/journal.pone.0105691. Consultado em 6 de dezembro de 2022 -
↑ Fu, Wenqing; O’Connor, Timothy D; Akey, Joshua M (1 de dezembro de 2013). «Genetic architecture of quantitative traits and complex diseases». Current Opinion in Genetics & Development. Genetics of system biology (em inglês) (6): 678–683. ISSN 0959-437X. doi:10.1016/j.gde.2013.10.008. Consultado em 11 de abril de 2022
-
↑ Fu, Wenqing; O’Connor, Timothy D.; Jun, Goo; Kang, Hyun Min; Abecasis, Goncalo; Leal, Suzanne M.; Gabriel, Stacey; Rieder, Mark J.; Altshuler, David (janeiro de 2013). «Analysis of 6,515 exomes reveals the recent origin of most human protein-coding variants». Nature (em inglês) (7431): 216–220. ISSN 1476-4687. doi:10.1038/nature11690. Consultado em 11 de abril de 2022
-
↑ NATHANIEL., JEANSON, (2022). TRACED;HUMAN DNA’S BIG SURPRISE. [S.l.]: MASTER BOOKS. OCLC 1306278031
-
↑ Wilson, James (julho de 2021). «The Genealogical Adam and Eve: The Surprising Science of Universal Ancestry». Bulletin for Biblical Research (2): 234–236. ISSN 1065-223X. doi:10.5325/bullbiblrese.31.2.0234. Consultado em 11 de abril de 2022
-
↑ JOSHUA., SWAMIDASS, S. (2021). GENEALOGICAL ADAM AND EVE : the surprising science of universal ancestry. [S.l.]: INTERVARSITY PRESS. OCLC 1253354060
-
↑ Kivisild, T.; Shen, P.; Wall, D.; Do, B.; Sung, R.; Davis, K.; Passarino, G.; Underhill, P.; Scharfe, C. (2006). «The Role of Selection in the Evolution of Human Mitochondrial Genomes». Genetics. doi:10.1534/genetics.105.043901. Consultado em 11 de abril de 2022
-
↑ Carter, R.; Lee, Stephen; Sanford, J. (2018). «An Overview of the Independent Histories of the Human Y Chromosome and the Human Mitochondrial chromosome». doi:10.15385/JPICC.2018.8.1.15. Consultado em 11 de abril de 2022
-
↑ Carter, R. (2007). «Mitochondrial diversity within modern human populations». Nucleic acids research. doi:10.1093/nar/gkm207. Consultado em 11 de abril de 2022
-
↑ Rohde, Douglas L. T.; Olson, Steve; Chang, Joseph T. (setembro de 2004). «Modelling the recent common ancestry of all living humans». Nature (em inglês) (7008): 562–566. ISSN 1476-4687. doi:10.1038/nature02842. Consultado em 11 de abril de 2022
-
↑ Sollars, Vincent; Lu, Xiangyi; Xiao, Li; Wang, Xiaoyan; Garfinkel, Mark D.; Ruden, Douglas M. (janeiro de 2003). «Evidence for an epigenetic mechanism by which Hsp90 acts as a capacitor for morphological evolution». Nature Genetics (em inglês) (1): 70–74. ISSN 1546-1718. doi:10.1038/ng1067. Consultado em 30 de julho de 2023
-
↑ Turner, Bryan M. (27 de novembro de 2009). «Epigenetic responses to environmental change and their evolutionary implications». Philosophical Transactions of the Royal Society B: Biological Sciences (em inglês) (1534): 3403–3418. ISSN 0962-8436. PMC 2781845
. PMID 19833651. doi:10.1098/rstb.2009.0125. Consultado em 30 de julho de 2023 -
↑ Kooke, Rik; Johannes, Frank; Wardenaar, René; Becker, Frank; Etcheverry, Mathilde; Colot, Vincent; Vreugdenhil, Dick; Keurentjes, Joost J.B. (1 de fevereiro de 2015). «Epigenetic Basis of Morphological Variation and Phenotypic Plasticity inArabidopsis thaliana». The Plant Cell (2): 337–348. ISSN 1532-298X. doi:10.1105/tpc.114.133025. Consultado em 30 de julho de 2023
-
↑ Vermeij, Geerat J. (15 de jan. de 2016). «Gigantism and Its Implications for the History of Life». PLOS ONE (em inglês) (1): e0146092. ISSN 1932-6203. PMC 4714876
. PMID 26771527. doi:10.1371/journal.pone.0146092. Consultado em 30 de julho de 2023 -
↑ Lamsdell, James C.; Braddy, Simon J. (23 de abril de 2010). «Cope’s Rule and Romer’s theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates». Biology Letters (em inglês) (2): 265–269. ISSN 1744-9561. PMC 2865068
. PMID 19828493. doi:10.1098/rsbl.2009.0700. Consultado em 30 de julho de 2023 -
↑ Braddy, Simon J; Poschmann, Markus; Tetlie, O. Erik (23 de fevereiro de 2008). «Giant claw reveals the largest ever arthropod». Biology Letters (em inglês) (1): 106–109. ISSN 1744-9561. PMC 2412931
. PMID 18029297. doi:10.1098/rsbl.2007.0491. Consultado em 30 de julho de 2023 -
↑ Schoenemann, Brigitte; Poschmann, Markus; Clarkson, Euan N. K. (28 de novembro de 2019). «Insights into the 400 million-year-old eyes of giant sea scorpions (Eurypterida) suggest the structure of Palaeozoic compound eyes». Scientific Reports (em inglês) (1). 17797 páginas. ISSN 2045-2322. doi:10.1038/s41598-019-53590-8. Consultado em 30 de julho de 2023
-
↑ Briggs, Derek E.G.; Roach, Brian T. (janeiro de 2020). «Excavating eurypterids, giant arthropods of the Palaeozoic». Geology Today (em inglês) (1): 16–21. ISSN 0266-6979. doi:10.1111/gto.12296. Consultado em 30 de julho de 2023
-
↑ Elvis, A. M.; Ekta, J. S. (2011). «Ozone therapy: A clinical review». Journal of Natural Science, Biology, and Medicine. 2 (1): 66–70. ISSN 0976-9668. PMC 3312702
. PMID 22470237. doi:10.4103/0976-9668.82319 -
↑ Hernandez, Alberto; Vinals, Montserrat; Pablos, Asuncion; Vilas, Francisco; Papadakos, Peter J.; Wijeysundera, Duminda N.; Vives, Marc (12 de junho de 2020). «Ozone therapy for patients with SARS-COV-2 pneumonia: a single-center prospective cohort study». medRxiv (em inglês): 2020.06.03.20117994. doi:10.1101/2020.06.03.20117994
-
↑ Schwartz, Adriana; Martínez-Sánchez, Gregorio; Lucía, Alejandra Menassa de; Viana, Sergio Mejía; Mita, Constanta Alina (12 de julho de 2020). «Complementary Application of the Ozonized Saline Solution in Mild and Severe Patients with Pneumonia Covid-19: A Non-randomized Pilot Study» (em inglês)
-
↑ Braidy, Nady; Izadi, Morteza; Sureda, Antoni; Jonaidi‐Jafari, Nematollah; Banki, Abdolali; Nabavi, Seyed F.; Nabavi, Seyed M. (2018). «Therapeutic relevance of ozone therapy in degenerative diseases: Focus on diabetes and spinal pain». Journal of Cellular Physiology (em inglês). 233 (4): 2705–2714. ISSN 1097-4652. doi:10.1002/jcp.26044
-
↑ Liu, Jian; Zhang, Peng; Tian, Jing; Li, Lun; Li, Jun; Tian, Jin Hui; Yang, KeHu (2015). «Ozone therapy for treating foot ulcers in people with diabetes». Cochrane Database of Systematic Reviews (em inglês) (10). ISSN 1465-1858. doi:10.1002/14651858.CD008474.pub2
-
↑ Zhang, Jing; Guan, Meiping; Xie, Cuihua; Luo, Xiangrong; Zhang, Qian; Xue, Yaoming (24 de junho de 2014). «Increased Growth Factors Play a Role in Wound Healing Promoted by Noninvasive Oxygen-Ozone Therapy in Diabetic Patients with Foot Ulcers». Oxidative Medicine and Cellular Longevity (em inglês). Consultado em 14 de agosto de 2020
-
↑ Tang, Wen-Juan; Jiang, Long; Wang, Ying; Kuang, Ze-Min (15 de dezembro de 2017). «Ozone therapy induced sinus arrest in a hypertensive patient with chronic kidney disease». Medicine. 96 (50). ISSN 0025-7974. PMC 5815785
. PMID 29390373. doi:10.1097/MD.0000000000009265 -
↑ Iovski, M. (2000). «Doppler blood flow velocity waveforms in hypertensive pregnant women». International Journal of Gynecology & Obstetrics. 70: D119–D119. ISSN 0020-7292. doi:10.1016/s0020-7292(00)84563-4
-
↑ Wang, Xiaoqi (28 de fevereiro de 2018). «Emerging roles of ozone in skin diseases». Zhong Nan Da Xue Xue Bao. Yi Xue Ban = Journal of Central South University. Medical Sciences. 43 (2): 114–123. ISSN 1672-7347. PMID 29559592. doi:10.11817/j.issn.1672-7347.2018.02.002
-
↑ Smetham, Graham P. (25 de julho de 2013). «Dawkins’ Darwinism Part I: Evolution – The Greatest Illusion on Earth and the New Quantum Platonic Paradigm». DNA Decipher Journal (em inglês) (3). ISSN 2159-046X. Consultado em 10 de abril de 2022
-
↑ Sakamoto, Wataru (27 de julho de 2009). «Faculty Opinions recommendation of Exchange of genetic material between cells in plant tissue grafts.». Faculty Opinions – Post-Publication Peer Review of the Biomedical Literature. Consultado em 10 de abril de 2022
-
↑ Williams, Brandon; Ahsan, Muhammad Umair; Frank, Margaret H (1 de fevereiro de 2021). «Getting to the root of grafting-induced traits». Current Opinion in Plant Biology. Growth and development 2021 (em inglês). 101988 páginas. ISSN 1369-5266. doi:10.1016/j.pbi.2020.101988. Consultado em 10 de abril de 2022
-
↑ Lönnig, Wolf-Ekkehard (2011). The Evolution of the Long-necked Giraffe (Giraffa Camelopardalis L.): What Do We Really Know? ; Testing the Theories of Gradualism, Macromutation, and Intelligent Design ; a Scientific Treatise (em inglês). [S.l.]: MV-Verlag
-
↑ Santos, Charles Morphy Dias dos (junho de 2008). «Os dinossauros de Hennig: sobre a importância do monofiletismo para a sistemática biológica». Scientiae Studia: 179–200. ISSN 1678-3166. doi:10.1590/S1678-31662008000200003. Consultado em 11 de abril de 2022
-
↑ Santos, Charles Morphy Dias dos (junho de 2008). «Os dinossauros de Hennig: sobre a importância do monofiletismo para a sistemática biológica». Scientiae Studia: 179–200. ISSN 1678-3166. doi:10.1590/S1678-31662008000200003. Consultado em 11 de abril de 2022
-
↑ Haber, Matthew H. (2019). «Species in the Age of Discordance». Philosophy, Theory, and Practice in Biology. ISSN 2475-3025. doi:10.3998/ptpbio.16039257.0011.021. Consultado em 11 de abril de 2022
-
↑ MICHAEL., BEHE, (2019). DARWIN DEVOLVES : why evolution has failed to explain how species progress and how science … shows it never will. [S.l.]: HARPER ONE. OCLC 1037282746
-
↑ Behe, Michael J. (31 de maio de 2013). «Theoretical Molecular Biology: Introductory Comments». WORLD SCIENTIFIC. doi:10.1142/9789814508728_others03. Consultado em 10 de abril de 2022
-
↑ author., Behe, Michael J., 1952-. Irreducible complexity. [S.l.: s.n.] OCLC 1181110519
-
↑ «James M. Tour». scholar.google.com. Consultado em 11 de abril de 2022
-
↑ Salisbury, Annis (14 de outubro de 1916). «The Tell-Tale Fossil». Scientific American (16): 348–349. ISSN 0036-8733. doi:10.1038/scientificamerican10141916-348. Consultado em 11 de abril de 2022
-
↑ Hetherington, Alexander J.; Berry, Christopher M.; Dolan, Liam (14 de junho de 2016). «Networks of highly branched stigmarian rootlets developed on the first giant trees». Proceedings of the National Academy of Sciences (em inglês) (24): 6695–6700. ISSN 0027-8424. PMC 4914198
. PMID 27226309. doi:10.1073/pnas.1514427113. Consultado em 11 de abril de 2022 -
↑ Lönnig, Wolf-Ekkehard; Saedler, Heinz (31 de dezembro de 1997). «Plant transposons: contributors to evolution?». Gene. Junk DNA: The Role and the Evolution of Non-Coding Sequences (em inglês) (1): 245–253. ISSN 0378-1119. doi:10.1016/S0378-1119(97)00397-1. Consultado em 11 de abril de 2022
-
↑ Scherer, Siegfried (21 de setembro de 1983). «Basic functional states in the evolution of light-driven cyclic electron transport». Journal of Theoretical Biology (em inglês) (2): 289–299. ISSN 0022-5193. doi:10.1016/0022-5193(83)90416-2. Consultado em 11 de abril de 2022
-
↑ Lambert, Grant R. (7 de abril de 1984). «Enzymic editing mechanisms and the origin of biological information transfer». Journal of Theoretical Biology (em inglês) (3): 387–403. ISSN 0022-5193. doi:10.1016/S0022-5193(84)80098-3. Consultado em 11 de abril de 2022
-
↑ Pollard, William G. (1 de outubro de 1984). «Rumors of transcendence in physics». American Journal of Physics: 877–881. ISSN 0002-9505. doi:10.1119/1.13901. Consultado em 11 de abril de 2022
-
↑ Kenyon, D. H. (1984). Matsuno, Koichiro; Dose, Klaus; Harada, Kaoru; Rohlfing, Duane L., eds. «A Comparison of Proteinoid and Aldocyanoin Microsystems as Models of the Primordial Protocell». Boston, MA: Springer US (em inglês): 163–188. ISBN 978-1-4684-4640-1. doi:10.1007/978-1-4684-4640-1_13. Consultado em 11 de abril de 2022
-
↑ Scherer, Siegfried (1 de dezembro de 1984). «Transmembrane electron transport and the neutral theory of evolution». Origins of life (em inglês) (1): 725–731. ISSN 1573-0875. doi:10.1007/BF00933727. Consultado em 11 de abril de 2022
-
↑ Dose, Klaus (1 de dezembro de 1988). «The Origin of Life: More Questions Than Answers». Interdisciplinary Science Reviews (4): 348–356. ISSN 0308-0188. doi:10.1179/isr.1988.13.4.348. Consultado em 11 de abril de 2022
-
↑ Axe, Douglas D (18 de agosto de 2000). «Extreme functional sensitivity to conservative amino acid changes on enzyme exteriors11Edited by J. Karn». Journal of Molecular Biology (em inglês) (3): 585–595. ISSN 0022-2836. doi:10.1006/jmbi.2000.3997. Consultado em 11 de abril de 2022
-
↑ C., Sanford, John (2008). Genetic entropy & the mystery of the genome. [S.l.]: FMS Publications. OCLC 422876474
-
↑ Wu, Haibo; Xing, Na; Meng, Kaiwen; Fu, Beibei; Xue, Weiwei; Dong, Pan; Tang, Wanyan; Xiao, Yang; Liu, Gexin (2021). «Nucleocapsid Mutations R203K/G204R Increase the Infectivity, Fitness and Virulence of SARS-CoV-2». SSRN Electronic Journal. ISSN 1556-5068. doi:10.2139/ssrn.3896432. Consultado em 11 de abril de 2022
-
↑ den Boer, P.J. (1 de junho de 1999). «Natural Selection or the Non-survival of the Non-fit». Acta Biotheoretica (em inglês) (2): 83–97. ISSN 1572-8358. doi:10.1023/A:1002053820381. Consultado em 11 de abril de 2022
-
↑ Falcon-Lang, Howard J.; Bashforth, Arden R. (1 de maio de 2004). «Pennsylvanian uplands were forested by giant cordaitalean trees». Geology (5): 417–420. ISSN 0091-7613. doi:10.1130/G20371.1. Consultado em 11 de abril de 2022
-
↑ Hetherington, Alexander J.; Berry, Christopher M.; Dolan, Liam (14 de junho de 2016). «Networks of highly branched stigmarian rootlets developed on the first giant trees». Proceedings of the National Academy of Sciences (em inglês) (24): 6695–6700. ISSN 0027-8424. PMC 4914198
. PMID 27226309. doi:10.1073/pnas.1514427113. Consultado em 11 de abril de 2022 -
↑ Donovan, S. K. (1 de maio de 2013). «Giant crinoid stems from the Lower Carboniferous (Mississippian) of Clitheroe, Lancashire, UK». Proceedings of the Yorkshire Geological Society (em inglês) (3): 211–218. ISSN 0044-0604. doi:10.1144/pygs2013-328. Consultado em 11 de abril de 2022
-
↑ Sanford, John C. (1 de dezembro de 1988). «The biolistic process». Trends in Biotechnology (em inglês) (12): 299–302. ISSN 0167-7799. doi:10.1016/0167-7799(88)90023-6. Consultado em 11 de abril de 2022
-
↑ Sanford, John C. (maio de 1990). «Biolistic plant transformation». Physiologia Plantarum (em inglês) (1): 206–209. ISSN 0031-9317. doi:10.1111/j.1399-3054.1990.tb05888.x. Consultado em 11 de abril de 2022
-
↑ Armaleo, Daniele; Ye, Guang-Ning; Klein, Theodore M.; Shark, Katherine B.; Sanford, John C.; Johnston, Stephen Albert (1 de fevereiro de 1990). «Biolistic nuclear transformation of Saccharomyces cerevisiae and other fungi». Current Genetics (em inglês) (2): 97–103. ISSN 1432-0983. doi:10.1007/BF00312852. Consultado em 11 de abril de 2022
-
↑ Sanford, John C. (setembro de 2000). «The development of the biolistic process». In Vitro Cellular & Developmental Biology – Plant (5): 303–308. ISSN 1054-5476. doi:10.1007/s11627-000-0056-9. Consultado em 11 de abril de 2022
-
↑ Shark, K B; Smith, F D; Harpending, P R; Rasmussen, J L; Sanford, J C (fevereiro de 1991). «Biolistic transformation of a procaryote, Bacillus megaterium». Applied and Environmental Microbiology (em inglês) (2): 480–485. ISSN 0099-2240. PMC 182736
. PMID 1901706. doi:10.1128/aem.57.2.480-485.1991. Consultado em 11 de abril de 2022 -
↑ Johnston, Stephen A.; Riedy, Mark; De Vit, Michael J.; Sanford, J. C.; McElligott, Sandra; Williams, R. Sanders (1 de janeiro de 1991). «Biolistic transformation of animal tissue». In Vitro Cellular & Developmental Biology – Plant (em inglês) (1): 11–14. ISSN 1475-2689. doi:10.1007/BF02632055. Consultado em 11 de abril de 2022
-
↑ Wise, Kurt (21 de outubro de 2020). «Baraminology: A Young-Earth Creation Biosystematic Method». Proceedings of the International Conference on Creationism (1). ISSN 2639-4006. Consultado em 11 de abril de 2022
-
↑ Sanders, Roger; Wise, Kurt (6 de outubro de 2020). «The Cognitum: A Perception-Dependent Concept Needed in Baraminology». Proceedings of the International Conference on Creationism (1). ISSN 2639-4006. Consultado em 11 de abril de 2022
-
↑ Brophy, Timothy (1 de janeiro de 2021). «A baraminological analysis of the landfowl (Aves: Galliformes)». Faculty Publications and Presentations. Consultado em 11 de abril de 2022
|
Esta é uma página de testes de Antigeocronologic, uma subpágina da principal. Serve como um local de testes e espaço de desenvolvimento, desta feita não é um artigo enciclopédico. Para uma página de testes sua, crie uma aqui. Como editar: Tutorial • Guia de edição • Livro de estilo • Referência rápida Como criar uma página: Guia passo a passo • Como criar • Verificabilidade • Critérios de notoriedade |
Efeitos de Grandes Impactos de Asteroides na Geologia Sedimentar, Geocronologia Radiométrica e DNA dos Seres Vivos
Sodré GB Neto
Resumo: Examinamos os efeitos catastróficos de impactos de asteroides na Terra, analisando suas consequências para a geologia sedimentar, a confiabilidade da geocronologia radiométrica e as mutações no DNA dos seres vivos por ocasião de grandes impactos de Asteroides e seus efeitos elétricos-radioativos. Apresentamos evidências científicas sobre como estes eventos extremos podem ter acelerado processos de decaimento radioativo, gerado formações geológicas extensas e causado alterações genéticas significativas nas espécies sobreviventes.
Introdução
Os impactos de grandes asteroides na Terra geraram fenômenos físicos extremos, incluindo atrito intenso, efeitos de spallação nuclear que produziram isótopos radioativos em rochas, efeitos piezoelétricos, temperaturas extremas instantâneas, ondas sonoras e diferenciais de carga que criaram plasmas de alta energia. A intensidade desses plasmas, caracterizados por altíssima amperagem, permitiu que elétrons ultrapassassem a barreira de Coulomb, provocando o decaimento acelerado de nêutrons e prótons tanto em elementos pesados quanto leves.
Esta pesquisa identifica, de maneira inédita, uma disparidade significativa no acúmulo de mutações ao longo da história dos seres vivos. Observamos um contraste entre o número limitado de mutações encontradas em múmias da antiguidade (aproximadamente 300 mutações mitocondriais) e o número extraordinariamente elevado de mutações na humanidade atual (cerca de 20.000 mapeadas pelo banco de dados MITOMAP). Esta disparidade não pode ser explicada pelas baixas taxas de acúmulo de mutações por geração (0,024 mutações mitocondriais por geração).
Argumentamos que a radiação gerada durante eventos de impacto, resultado da ultrapassagem da barreira de Coulomb em condições de plasma, prejudicou o DNA dos organismos sobreviventes, gerando incontáveis mutações, principalmente do tipo SNPs (polimorfismos de nucleotídeo único). Ademais, os plasmas de alta energia gerados por estes impactos invalidaram o pressuposto fundamental da constância do decaimento radioativo que fundamenta as datações radiométricas convencionais.
Introdução à Física de Impactos de Asteroides
A interação da Terra com corpos celestes, especialmente asteroides, tem sido uma força determinante na modelagem da superfície terrestre e na influência sobre a evolução da vida ao longo do tempo geológico. A análise de impactos de asteroides e suas consequências é fundamental para a compreensão da história geológica do planeta e das condições ambientais que permitiram ou limitaram o desenvolvimento da vida.
Os impactos de asteroides não são eventos simples de colisão, mas fenômenos extremamente complexos que envolvem a liberação repentina de quantidades extraordinárias de energia. Quando um asteroide colide com a Terra, desencadeia-se uma série de processos físicos e químicos que alteram significativamente a geologia local e, dependendo da magnitude do impacto, podem ter efeitos globais. A temperatura, pressão e outras condições extremas geradas no momento do impacto criam um ambiente único que não é replicado em nenhum outro processo natural terrestre.
A integração de dados multidisciplinares é essencial para compreender completamente os impactos de asteroides e suas consequências. Estudos geológicos fornecem evidências diretas dos efeitos físicos dos impactos, enquanto análises geoquímicas e isotópicas revelam alterações na composição das rochas afetadas. Por outro lado, estudos biológicos e paleontológicos oferecem insights sobre como esses eventos afetaram a evolução e distribuição dos seres vivos.
Evidências Geológicas de Impactos na Terra
As evidências geológicas de impactos de asteroides na Terra são diversas e frequentemente dramáticas. Entre as mais importantes estão as estruturas de impacto, que variam desde pequenas crateras até enormes bacias de impacto com centenas de quilômetros de diâmetro. Estas estruturas apresentam características distintivas, como bordas elevadas, bacias centrais e padrões de fratura radial e concêntrica, que as diferenciam de outras formações geológicas.
A análise de Schmitz e Bowring (2001) demonstra como os impactos extraterrestres influenciaram profundamente a evolução geológica do planeta. Seus estudos revelam que eventos de impacto significativos coincidiram com importantes transições no registro geológico, sugerindo uma relação causal entre impactos e mudanças geológicas de grande escala. Reimold e Gibson (1996) fornecem uma revisão abrangente das evidências geológicas de crateras de impacto, descrevendo as características diagnósticas que permitem identificar estruturas de impacto mesmo quando altamente degradadas pela erosão.
Bottke et al. (2002) discutem as origens dos asteroides e suas implicações para a frequência e intensidade de chuvas de impactos ao longo da história da Terra. Seus modelos sugerem que a Terra foi submetida a períodos de bombardeamento intensificado, possivelmente relacionados a eventos de disrupção de asteroides no cinturão principal ou a perturbações na nuvem de Oort. Estes períodos de aumento na frequência de impactos podem ter tido consequências profundas para a evolução da crosta terrestre e para os processos sedimentares.
Atrito e Geração de Calor Durante Impactos
O atrito gerado durante o impacto de um asteroide representa um dos aspectos mais energéticos desse fenômeno. Quando um corpo celeste atinge a superfície terrestre a velocidades hipersônicas, tipicamente entre 15 e 30 km/s, a fricção resultante da interação entre o projétil e o material alvo produz um aquecimento extremo, que pode atingir temperaturas superiores a 10.000°C em questão de milissegundos.
Este processo de aquecimento não se limita apenas ao ponto de impacto. A energia térmica se propaga radialmente através do solo, criando zonas concêntricas de metamorfismo térmico. Nas regiões mais próximas ao epicentro, o calor é suficiente para vaporizar instantaneamente rochas e minerais, transformando-os em um plasma de alta temperatura. Em zonas intermediárias, ocorre a fusão parcial ou total do material rochoso, enquanto áreas mais distantes experimentam recristalização e outras alterações mineralógicas devido ao choque térmico.
De acordo com os estudos de Zhang et al. (2008), esse atrito extremo também contribui para a aceleração de elétrons a altas energias, criando condições para reações nucleares nas rochas impactadas. O calor gerado pelo atrito provoca a excitação de elétrons nos átomos, resultando em ionização e, em casos extremos, na quebra de ligações nucleares. Os efeitos térmicos do impacto persistem por períodos variáveis, dependendo da magnitude do evento. Grandes impactos podem criar anomalias térmicas que permanecem por décadas ou até séculos, alterando significativamente os padrões climáticos regionais e globais.
Temperatura Imediata e Efeitos Térmicos de Impactos
O aumento instantâneo de temperatura representa um dos aspectos mais devastadores dos impactos de asteroides. No momento do impacto, a energia cinética do asteroide é convertida principalmente em energia térmica, gerando temperaturas que podem exceder dezenas de milhares de graus Celsius no ponto de colisão – valores comparáveis à superfície do Sol, conforme demonstrado por Collins et al. (2005) e Wünnemann et al. (2008).
Este calor extremo vaporiza instantaneamente tanto o asteroide quanto as rochas no ponto de impacto, criando uma nuvem de vapor superaquecido que se expande rapidamente. O material rochoso vaporizado pode alcançar temperaturas de 8.000 a 10.000°C, formando uma pluma ascendente que se eleva na atmosfera, como descrito por Artemieva & Morgan (2009) e Johnson & Melosh (2012). Quando este material resfria e se condensa, pode precipitar como pequenas esferas de vidro (microtectitos) ou fragmentos angulares que são distribuídos globalmente em eventos de grande magnitude, conforme documentado por Glass & Simonson (2013).
A radiação térmica emitida pela pluma e pelos materiais ejetados pode causar incêndios em áreas extremamente distantes do ponto de impacto. No caso do impacto de Chicxulub, evidências sugerem que incêndios florestais em escala global foram desencadeados pela radiação térmica intensa que atingiu a superfície terrestre quando os fragmentos ejetados reentram na atmosfera, criando um fenômeno conhecido como “chuva de meteoros secundária”, segundo Robertson et al. (2013) e Bardeen et al. (2017).
Consequências Ambientais Globais de Grandes Impactos
Os impactos de asteroides de grande magnitude produzem consequências ambientais que transcendem significativamente os efeitos locais. Argumenta-se que estes eventos são excepcionalmente mais letais em escala global do que quaisquer outras causas terrestres propostas para extinções em massa, devido a duas características únicas: a instantaneidade dos efeitos ambientais e suas consequências compostas.
A rapidez com que as mudanças ambientais ocorrem após um impacto – em escalas de tempo de horas a meses – não permite que as espécies desenvolvam adaptações evolutivas ou migrem para áreas de refúgio. Essa velocidade de alteração ambiental é incomparável a qualquer outro fenômeno natural, incluindo erupções vulcânicas ou mudanças climáticas graduais. Em questão de horas, ecossistemas inteiros podem colapsar devido às condições extremas geradas pelo impacto.
Além da rapidez, a natureza composta dos efeitos ambientais torna os impactos particularmente devastadores. Um único evento desencadeia múltiplos estressores ambientais: céus superaquecidos durante a reentrada dos ejecta na atmosfera, incêndios globais, destruição da camada de ozônio, terremotos e tsunamis, seguidos por meses de “inverno de impacto” e séculos de aquecimento global, além do envenenamento dos oceanos. Esta combinação sinérgica de efeitos cria condições quase impossíveis para a sobrevivência de muitas espécies, especialmente as mais especializadas ou de maior porte.
Vantagens Letais dos Impactos Sobre Outros Eventos Catastróficos
Quando comparados a outros eventos catastróficos como vulcanismo intenso, regressões marinhas ou colapsos hipotéticos de plataformas continentais e calotas polares, os impactos de asteroides apresentam um potencial destrutivo substancialmente maior. Esta diferença fundamenta-se principalmente na abruptez dos efeitos e na impossibilidade de adaptação evolutiva em tempo hábil.
O vulcanismo, mesmo em seus episódios mais intensos como as grandes províncias ígneas, libera gases e materiais ao longo de milhares a milhões de anos, permitindo certa adaptação gradual dos ecossistemas. De forma similar, as regressões marinhas, ainda que significativas, ocorrem em escalas de tempo geológicas que possibilitam migrações e adaptações das comunidades biológicas. Em contraste, os impactos de asteroides provocam alterações ambientais drásticas em questão de horas ou dias, eliminando virtualmente qualquer possibilidade de resposta adaptativa.
As formas de vida têm oportunidades consideravelmente melhores de sobrevivência em cenários de duração mais longa, onde mecanismos de refúgio ecológico, migração para habitats favoráveis ou adaptação evolutiva podem operar. A natureza instantânea e global dos efeitos de impacto elimina estas rotas de sobrevivência, resultando em taxas de extinção potencialmente muito mais elevadas. Esta característica única dos impactos de asteroides sugere que estes eventos, mais do que quaisquer outros, podem ter sido os principais responsáveis pelas grandes extinções em massa registradas no registro fóssil.
Plausibilidade dos Impactos como Causadores de Extinções em Massa
Considerando a inevitabilidade histórica de impactos de Objetos Próximos à Terra (NEA – Near Earth Asteroids) e sua capacidade destrutiva sem paralelos, é altamente plausível que estes eventos tenham sido os principais responsáveis pelas extinções em massa ao longo da história geológica. Esta hipótese, inicialmente proposta por David Raup, ganha força quando analisamos a eficácia incomparável dos impactos em gerar condições ambientais extremas e globalmente letais.
Mesmo na ausência de evidências diretas para extinções específicas, a questão fundamental permanece: que outro processo natural poderia ser tão devastadoramente eficaz? A combinação única de efeitos instantâneos e sinérgicos torna os impactos de asteroides e cometas candidatos primários para explicar os principais eventos de extinção registrados no registro fóssil. Além disso, mesmo que algumas extinções tenham tido outras causas primárias, é virtualmente impossível que os maiores impactos ocorridos durante o Fanerozoico não tenham deixado marcas significativas na biodiversidade global.
Esta perspectiva sugere que o registro fóssil, com suas descontinuidades abruptas na diversidade biológica, pode refletir primariamente a história dos grandes impactos terrestres, em vez de processos graduais internos ao planeta. As extinções em massa, vistas por este prisma, seriam testemunhos não de lentas mudanças ambientais, mas de catástrofes cósmicas que repetidamente redesenharam o curso da evolução biológica na Terra.
Processos de Fusão Nuclear em Impactos
Um dos aspectos mais controversos e fascinantes da física de impactos de asteroides é a possibilidade de ocorrência de processos de fusão nuclear em pequena escala. A fusão nuclear, o mesmo processo que alimenta as estrelas, requer condições extremas de temperatura e pressão para superar a repulsão eletrostática entre núcleos atômicos e permitir que se fundam, liberando energia, conforme destacado nos trabalhos de Crawford & Schultz (2014) e Boslough & Crawford (2008).
Durante o impacto de grandes asteroides, as temperaturas no ponto de colisão podem atingir dezenas de milhares de graus Celsius, aproximando-se das condições encontradas na superfície do Sol. Simultaneamente, as pressões instantâneas podem exceder milhões de atmosferas, como demonstrado por Melosh & Collins (2019) e Pierazzo & Artemieva (2012). Nestas condições extremas, particularmente no plasma de alta energia gerado pelo impacto, íons de elementos leves como hidrogênio, deutério e trítio podem ocasionalmente se aproximar o suficiente para que a força nuclear forte supere a repulsão eletrostática, resultando em fusão, conforme sugerido por Svetsov & Shuvalov (2016) e Tagle & Hecht (2006).
Evidências indiretas de possíveis processos de fusão durante impactos podem ser encontradas na análise de isótopos anômalos em rochas impactadas. Por exemplo, concentrações incomuns de hélio-3, um produto típico de certas reações de fusão, têm sido identificadas em vidros de impacto (tectitos), como reportado por Koeberl et al. (2018) e Simonson & Glass (2004). Além disso, a presença de elementos leves com razões isotópicas alteradas poderia ser explicada por processos limitados de fusão nuclear, conforme proposto por Qin & Humayun (2020) e Jourdan et al. (2012).
Extensão e Significado das Reações de Fusão em Impactos
É fundamental ressaltar que, se ocorrer, a fusão nuclear durante impactos de asteroides seria um fenômeno localizado e de curta duração, não comparável em escala às reações contínuas que ocorrem no interior do Sol, como apontado por Johnson & Melosh (2022) e French & Koeberl (2010). Estas reações ocorreriam principalmente nas regiões mais energéticas do plasma gerado pelo impacto e teriam duração limitada ao período em que condições extremas de temperatura e pressão são mantidas.
Apesar de sua escala limitada, mesmo processos restritos de fusão nuclear contribuiriam significativamente para o inventário total de energia liberada durante o impacto. Mais importante ainda, estas reações poderiam produzir assinaturas geoquímicas distintas que auxiliam os cientistas na identificação de antigos locais de impacto, mesmo quando outras evidências morfológicas foram obliteradas pela erosão e tectônica, como sugerem Glass & Simonson (2017) e Reimold & Koeberl (2014).
Do ponto de vista da geocronologia, a ocorrência de reações de fusão nuclear durante impactos tem implicações profundas para os métodos de datação radiométrica. Estas reações poderiam alterar localmente as proporções isotópicas utilizadas em datações, potencialmente levando a interpretações errôneas de idades absolutas. Esta possibilidade sugere a necessidade de cautela ao interpretar idades radiométricas de rochas em regiões conhecidas por terem sofrido impactos significativos, uma consideração raramente incorporada nos protocolos convencionais de datação geológica.
Efeito Piezoelétrico em Rochas Impactadas
O efeito piezoelétrico, embora frequentemente associado a cristais como quartzo em aplicações tecnológicas, desempenha um papel fundamental durante impactos de asteroides. Este fenômeno ocorre quando certos minerais, principalmente silicatos como quartzo e feldspato, geram uma diferença de potencial elétrico em resposta à deformação mecânica extrema causada pelo impacto, como demonstrado por Wang et al. (2016).
Quando as ondas de choque do impacto se propagam através da crosta terrestre, exercem pressões instantâneas enormes sobre os cristais rochosos, que podem exceder centenas de gigapascais. Nos minerais piezoelétricos, essa compressão força um realinhamento das cargas elétricas internas, criando momentaneamente campos elétricos localizados de alta intensidade. Em rochas ricas em quartzo, como granitos e arenitos, esse efeito pode ser particularmente pronunciado, gerando diferenças de potencial da ordem de milhares de volts em escalas microscópicas.
Esses campos elétricos transitórios contribuem significativamente para a ionização do ar e dos materiais vaporizados, facilitando a formação de plasmas. Além disso, podem interagir com os campos magnéticos gerados pela movimentação de material condutor durante o impacto, criando complexas interações eletromagnéticas. O efeito piezoelétrico também pode acelerar partículas carregadas, especialmente elétrons, amplificando os processos de spallação e potencialmente contribuindo para reações nucleares induzidas por impacto, fenômenos que têm implicações diretas para a confiabilidade das datações radiométricas em rochas afetadas.
Implicações Geoquímicas do Efeito Piezoelétrico
As implicações do efeito piezoelétrico durante impactos vão muito além da física imediata do evento. Os campos elétricos gerados piezoeletricamente podem induzir reações químicas não convencionais nas rochas impactadas, contribuindo para a formação de minerais e compostos que normalmente não se formariam em condições geológicas padrão. Esta alteração na composição mineralógica e química das rochas impactadas cria assinaturas geoquímicas distintas que podem persistir por bilhões de anos.
Um dos aspectos mais significativos dessas anomalias mineralógicas é seu valor como indicadores diagnósticos de antigos locais de impacto. Mesmo quando a morfologia original da cratera foi completamente erodida ou deformada por processos tectônicos subsequentes, a presença de minerais de alta pressão, vidros de impacto com composições químicas anômalas e estruturas de deformação planar em cristais de quartzo permite aos geólogos identificar inequivocamente o local como tendo sofrido um impacto de asteroide no passado.
Do ponto de vista da geocronologia, os campos elétricos piezoelétricos podem potencialmente alterar as taxas de decaimento radioativo em escala local, acelerando temporariamente processos que são assumidos como constantes nos métodos convencionais de datação radiométrica. Esta possibilidade levanta questões importantes sobre a confiabilidade das idades absolutas obtidas para rochas que estiveram sujeitas a impactos significativos, sugerindo a necessidade de reavaliação de alguns princípios fundamentais da geocronologia em contextos de eventos de alta energia.
Formação de Plasma em Grandes Impactos
Um dos fenômenos mais espetaculares e energéticos resultantes do impacto de grandes asteroides é a formação de plasma – um estado da matéria altamente ionizado composto por elétrons livres e íons positivos. Este quarto estado da matéria se forma quando temperaturas extremas e campos elétricos intensos provocam a separação dos elétrons de seus átomos, criando um gás condutor que pode interagir fortemente com campos eletromagnéticos, conforme descrito por Lieberman & Lichtenberg (2005).
Nos primeiros instantes após o impacto, a combinação de temperaturas que podem exceder dezenas de milhares de graus Celsius, campos elétricos gerados por efeitos piezoelétricos e a intensa pressão da onda de choque criam condições ideais para a ionização em massa do material vaporizado. O plasma resultante pode se estender por vários quilômetros acima do ponto de impacto, formando uma coluna luminosa visível a grandes distâncias, semelhante a um relâmpago gigantesco mas muito mais energético e persistente.
A física deste plasma de impacto é extremamente complexa. Devido à alta amperagem – que pode atingir milhões de amperes – correntes elétricas massivas fluem através do plasma, gerando campos magnéticos intensos. Estes campos, por sua vez, podem confinar e direcionar o plasma, criando estruturas filamentares e vórtices. Relâmpagos gigantescos podem ser observados nessa fase, como resultado das diferenças de potencial elétrico e da alta condutividade do meio ionizado, fenômenos que contribuem para a complexa cadeia de eventos que caracteriza a física de impactos de alta energia.
Aceleração de Elétrons no Plasma de Impacto
Um aspecto particularmente significativo da formação de plasma durante impactos de asteroides é que, no interior deste meio ionizado, elétrons podem ser acelerados a velocidades relativísticas. Conforme destacado por McCoy et al. (2013) e Zhang et al. (2008), estas partículas energéticas podem atingir energias suficientes para superar a barreira de Coulomb – a força de repulsão eletrostática entre partículas de mesma carga – permitindo interações com núcleos atômicos que normalmente seriam energeticamente desfavoráveis.
A aceleração de elétrons no plasma ocorre principalmente através de dois mecanismos: aceleração por campo elétrico direto e aceleração estocástica em turbulências do plasma. No primeiro caso, os intensos campos elétricos gerados pela separação de cargas no plasma exercem força direta sobre os elétrons, acelerando-os a energias da ordem de MeV ou até GeV em casos extremos. No segundo mecanismo, elétrons ganham energia através de múltiplas interações com ondas de plasma turbulento, um processo análogo à aceleração de partículas cósmicas em choques astrofísicos.
Estes elétrons de alta energia desempenham papel crucial em diversos processos secundários associados ao impacto. Eles facilitam tanto a spallação nuclear – onde núcleos atômicos são fragmentados pela colisão com partículas energéticas – quanto, potencialmente, processos de fusão nuclear em pequena escala. Além disso, a radiação de freamento (bremsstrahlung) emitida por estes elétrons ao serem desacelerados gera raios X e gama que contribuem para o intenso campo de radiação associado ao evento de impacto, com consequências significativas para sistemas biológicos expostos.
Spallação Nuclear em Impactos de Asteroides
A spallação nuclear representa um dos fenômenos mais fascinantes e menos compreendidos associados aos impactos de asteroides. Este processo ocorre quando partículas de alta energia, geradas durante o impacto, colidem com núcleos atômicos nas rochas, fragmentando-os e liberando nêutrons, prótons e partículas alfa. O resultado é a produção de isótopos radioativos que normalmente não existiriam em abundância na crosta terrestre.
Durante um impacto de alta energia, os elétrons são acelerados a velocidades relativísticas devido ao imenso campo eletromagnético gerado. Esses elétrons energéticos, ao interagirem com os núcleos dos átomos presentes nas rochas, desencadeiam reações nucleares que alteram a composição isotópica dos elementos. Conforme indicado por Zhang et al. (2008), essa aceleração de elétrons durante impactos de asteroides pode atingir energias suficientes para induzir reações nucleares significativas, criando um laboratório natural de física nuclear de alta energia.
Os isótopos radioativos formados por spallação funcionam como “relógios geológicos”, permitindo aos cientistas datar eventos de impacto com precisão considerável. Elementos como berílio-10, alumínio-26 e cloro-36 são particularmente importantes nesse contexto, pois suas meias-vidas são conhecidas e sua presença anômala em rochas pode indicar exposição a eventos de spallação. Além de seu valor como marcadores temporais, os isótopos radioativos produzidos por spallação também contribuem para o aumento da radiação local após o impacto, um fator adicional de estresse para os organismos sobreviventes.
Superação da Barreira de Coulomb
A barreira de Coulomb representa um dos princípios fundamentais da física nuclear, consistindo na força de repulsão eletrostática que impede que núcleos atômicos com cargas positivas se aproximem o suficiente para que ocorram reações nucleares. Em condições normais, esta barreira atua como um escudo protetor que mantém a estabilidade dos átomos, exigindo energias extremamente altas para ser superada, conforme explicado por Bertsch et al. (2014).
Durante o impacto de grandes asteroides, no entanto, condições extraordinárias permitem que esta barreira seja temporariamente vencida. Os elétrons acelerados no plasma de alta energia gerado pelo impacto podem atingir velocidades próximas à da luz. Quando estes elétrons relativísticos colidem com núcleos atômicos, podem transferir energia suficiente para comprimir temporariamente a nuvem eletrônica, reduzindo efetivamente a distância entre núcleos vizinhos e facilitando interações que normalmente seriam impedidas pela repulsão eletrostática.
Além disso, as altíssimas temperaturas e pressões resultantes do impacto fornecem energia térmica adicional aos núcleos, aumentando a probabilidade de tunelamento quântico através da barreira de Coulomb. Este fenômeno, conhecido como efeito de tunelamento, permite que partículas com energia insuficiente para superar uma barreira energética ainda assim consigam atravessá-la, graças aos princípios da mecânica quântica. Estes mecanismos combinados criam condições para reações nucleares que normalmente só ocorreriam em ambientes extremos como o interior de estrelas.
Implicações da Superação da Barreira de Coulomb
A superação da barreira de Coulomb em ambientes de impacto tem implicações profundas para a geoquímica das rochas afetadas e para nossa compreensão dos processos de datação radiométrica. Este fenômeno permite o decaimento acelerado de isótopos instáveis e facilita reações de transmutação nuclear, onde um elemento pode ser convertido em outro através de processos nucleares induzidos pelo impacto.
Estas transformações nucleares contribuem para a formação de isótopos raros e elementos que normalmente não seriam encontrados nas concentrações observadas em rochas impactadas, fornecendo uma assinatura geoquímica única desses eventos catastróficos. Por exemplo, anomalias nas concentrações de isótopos de gases nobres como hélio, neônio e argônio, frequentemente observadas em tectitos e outros materiais de impacto, podem ser explicadas por reações nucleares facilitadas pela superação da barreira de Coulomb.
Do ponto de vista da geocronologia, este fenômeno levanta questões fundamentais sobre a confiabilidade dos métodos de datação radiométrica em rochas que foram submetidas a impactos significativos. Se a superação da barreira de Coulomb durante impactos permite o decaimento acelerado de isótopos, então a premissa fundamental de taxas de decaimento constantes, que sustenta todos os métodos de datação radiométrica, pode ser violada em contextos de impacto. Esta possibilidade sugere que idades radiométricas obtidas de rochas impactadas podem refletir não o tempo real desde a formação da rocha, mas sim efeitos de “envelhecimento aparente” causados por processos nucleares acelerados durante o evento de impacto.
Decaimento Acelerado de Nêutrons e Prótons
Um dos fenômenos mais extraordinários associados aos impactos de grandes asteroides é o decaimento acelerado de partículas subatômicas, particularmente nêutrons e prótons. Em condições normais, prótons são extremamente estáveis (com meia-vida teórica superior à idade do universo), enquanto nêutrons livres têm uma meia-vida de aproximadamente 15 minutos antes de decair em um próton, um elétron e um antineutrino.
No ambiente de alta energia criado por um impacto de asteroide, as regras convencionais da física nuclear são temporariamente alteradas. As intensas forças eletromagnéticas geradas no plasma de impacto podem desestabilizar partículas subatômicas, tanto em elementos leves quanto pesados. Nêutrons podem ser ejetados dos núcleos através de reações de spallação e, uma vez livres, seu decaimento pode ser significativamente acelerado pelas condições extremas presentes, produzindo fluxos intensos de radiação beta e neutrinos.
Esse decaimento acelerado tem várias consequências importantes. Primeiro, contribui para a liberação adicional de energia na forma de radiação beta (elétrons de alta energia) e raios gama. Segundo, altera a composição isotópica das rochas impactadas, criando razões isotópicas anômalas que podem ser detectadas mesmo bilhões de anos após o evento. Terceiro, a transmutação nuclear resultante pode produzir elementos e isótopos raros, alguns dos quais radioativos com meias-vidas variáveis, criando uma “impressão digital” nuclear única do evento de impacto.
Evidências do Decaimento Acelerado em Rochas Impactadas
As evidências do processo de decaimento acelerado de partículas subatômicas podem ser encontradas na análise detalhada de rochas impactadas. Concentrações anormais de certos isótopos, como hélio-3, berílio-10 ou neônio-21, são frequentemente interpretadas como evidências de reações nucleares induzidas por impacto. Estas anomalias isotópicas constituem uma “impressão digital” nuclear que permite aos geocientistas identificar e datar antigos eventos de impacto, mesmo quando outras evidências morfológicas já foram erodidas pelo tempo.
Estudos mineralógicos detalhados de materiais impactados revelam frequentemente a presença de minerais exóticos e fases metaestáveis que só podem se formar sob condições extremas de temperatura e pressão. A presença destes minerais, como a stishovita (uma forma de alta pressão de sílica) ou a coesita, é consistente com os ambientes de alta energia necessários para induzir o decaimento acelerado de partículas subatômicas. Além disso, análises de microtexturas em minerais impactados frequentemente mostram evidências de danos por radiação consistentes com a exposição a fluxos intensos de partículas subatômicas durante o evento de impacto.
Do ponto de vista da datação, estas evidências sugerem que os métodos radiométricos convencionais podem fornecer resultados enganosos quando aplicados a rochas que sofreram impactos significativos. Se o decaimento de isótopos foi temporariamente acelerado durante o evento de impacto, as idades calculadas podem parecer muito mais antigas do que a idade real da rocha. Esta possibilidade exige uma reavaliação cuidadosa das cronologias geológicas estabelecidas, especialmente em regiões conhecidas por terem experimentado eventos de impacto significativos.
Emissão de Radiação Durante Impactos
A emissão de radiação durante eventos de impacto de asteroides representa um aspecto crítico tanto para a compreensão da física desses fenômenos quanto para a avaliação de seus efeitos biológicos. Quando um grande asteroide colide com a Terra, múltiplos mecanismos contribuem para a liberação de diferentes tipos de radiação ionizante e não ionizante, criando um ambiente temporariamente hostil à vida em vastas áreas ao redor do ponto de impacto.
A radiação térmica constitui a primeira e mais óbvia forma de emissão. O calor intenso gerado pelo impacto produz radiação infravermelha e luz visível em quantidades massivas, potencialmente causando incêndios em áreas distantes do epicentro. Para impactos verdadeiramente grandes, como o evento K-T de 65 milhões de anos atrás, estima-se que a radiação térmica tenha sido suficiente para aquecer a atmosfera global a temperaturas próximas de 100°C por várias horas, criando condições incompatíveis com a vida na superfície terrestre.
A radiação ionizante, incluindo raios X, raios gama e partículas de alta energia (prótons, nêutrons e elétrons), é produzida através de vários processos nucleares já mencionados: spallação, decaimento acelerado e, em casos extremos, possíveis reações de fusão em pequena escala dentro do plasma de impacto. Essa radiação ionizante penetra profundamente em materiais orgânicos, danificando DNA e proteínas, e pode ser particularmente letal para organismos complexos que não possuem mecanismos eficientes de reparo de danos radioativos.
Efeitos Biológicos da Radiação de Impacto
Os efeitos biológicos da radiação gerada por impactos de asteroides são potencialmente devastadores para a biosfera. A radiação ionizante, em particular, interage diretamente com o material genético dos organismos, causando quebras nas cadeias de DNA e produzindo radicais livres que danificam estruturas celulares. A intensidade da radiação liberada durante grandes impactos pode exceder em muitas ordens de magnitude os níveis considerados letais para a maioria das formas de vida.
Organismos complexos, como animais vertebrados, são especialmente vulneráveis aos efeitos da radiação ionizante. Mesmo doses relativamente baixas podem causar síndrome aguda de radiação, caracterizada por náuseas, vômitos, hemorragias e eventualmente morte. Doses mais altas, como as esperadas nas proximidades de um grande impacto, causariam morte celular generalizada e falência de órgãos em questão de horas. Para organismos menores e mais simples, como bactérias e arqueias, a resistência à radiação é geralmente maior, mas ainda assim existe um limite superior além do qual mesmo estes microorganismos não conseguem sobreviver.
Além dos efeitos agudos, a radiação também pode causar danos genéticos permanentes que afetam gerações futuras. Mutações induzidas por radiação são frequentemente deletérias e podem levar a um aumento na incidência de malformações congênitas, câncer e redução da fertilidade nas populações sobreviventes. Estes efeitos de longo prazo podem continuar a influenciar a evolução das espécies sobreviventes por muitas gerações após o impacto, criando pressões seletivas adicionais que moldam a recuperação da biodiversidade após o evento catastrófico.
Novos Modelos Sobre a Formação do Manto Terrestre
Novos modelos sobre a formação e dinâmica do manto terrestre têm sido propostos por equipes de geofísicos, incluindo trabalhos notáveis liderados por John Baumgardner. Estas pesquisas oferecem perspectivas alternativas sobre processos geológicos fundamentais e questionam alguns aspectos dos modelos convencionais de evolução da Terra. Baumgardner e colaboradores têm desenvolvido modelos computacionais sofisticados que simulam o comportamento do manto terrestre sob diferentes condições, permitindo novas interpretações de dados geofísicos.
Um exemplo significativo desses avanços é o trabalho publicado por Cho, Horstemeyer e Baumgardner (2017), que apresenta um modelo unificado estático e dinâmico de recristalização para os minerais do manto terrestre utilizando variáveis de estado interno. Este modelo representa uma abordagem inovadora para compreender como o manto responde a tensões e deformações, integrando processos que ocorrem em escalas temporais e espaciais muito diferentes. A implementação deste modelo permite simulações mais precisas do comportamento do manto terrestre durante eventos catastróficos de alta energia, como impactos de asteroides ou rápidos movimentos tectônicos.
Estas novas abordagens para a modelagem do manto terrestre são particularmente relevantes quando consideramos os efeitos de grandes impactos de asteroides. Impactos de magnitude suficiente podem perturbar significativamente a estrutura e dinâmica do manto, potencialmente desencadeando episódios de atividade tectônica acelerada, vulcanismo intensificado e outros fenômenos geológicos de larga escala. Os modelos desenvolvidos por Baumgardner e colegas oferecem ferramentas para explorar estas possibilidades com maior rigor matemático e físico.
Questionamentos aos Métodos de Datação Absoluta
Além de desenvolver novos modelos para a dinâmica do manto terrestre, John Baumgardner e outros pesquisadores têm questionado aspectos fundamentais dos métodos de datação radiométrica absoluta. Um dos argumentos mais intrigantes apresentados por estes cientistas baseia-se na detecção inesperada de carbono-14 em materiais que, segundo a geocronologia convencional, deveriam ser antigos demais para conter este isótopo radioativo.
O carbono-14 possui uma meia-vida relativamente curta de aproximadamente 5.730 anos, o que significa que após cerca de 50.000 a 70.000 anos, a quantidade de carbono-14 remanescente em uma amostra deveria ser tão pequena que se tornaria indetectável pelos métodos analíticos atuais. No entanto, Baumgardner e sua equipe publicaram em 2004 resultados surpreendentes de análises realizadas no laboratório de Los Álamos, demonstrando a presença de quantidades mensuráveis de carbono-14 em amostras de material orgânico incrustado em rochas do Fanerozoico, consideradas como tendo entre 300 e 500 milhões de anos de idade segundo métodos convencionais.
Mais significativamente, estes pesquisadores também detectaram carbono-14 em diamantes, que são particularmente resistentes à contaminação devido à sua estrutura cristalina extremamente estável e densa. De acordo com a interpretação destes cientistas, a presença de carbono-14 em diamantes supostamente antigos sugere que estas rochas não podem ter a idade de centenas de milhões de anos atribuída a elas pela geocronologia convencional, mas devem ser significativamente mais recentes – não mais antigas que algumas dezenas de milhares de anos. Esta observação levanta questões fundamentais sobre a confiabilidade dos métodos de datação absoluta e suas premissas subjacentes.
Evidências de Catástrofes Geológicas Recentes
A superfície terrestre apresenta numerosos indícios de eventos catastróficos de grande magnitude ocorridos em um passado geologicamente recente. Estas evidências abrangem desde características geomorfológicas até anomalias sedimentológicas e geoquímicas que desafiam interpretações baseadas em processos graduais e uniformes ao longo de vastos períodos de tempo.
Uma categoria significativa dessas evidências inclui as vastas formações sedimentares que se estendem desde o período Ediacarano-Cambriano até o Pleistoceno. Estas camadas, frequentemente caracterizadas por sua notável continuidade lateral e espessura substancial, apresentam características texturais e sedimentológicas que sugerem deposição rápida em condições de alta energia, possivelmente relacionadas a eventos catastróficos como tsunamis gigantescos desencadeados por impactos de asteroides. A presença de fósseis bem preservados, muitas vezes em posição de vida ou mostrando evidências de sepultamento rápido, corrobora esta interpretação.
Outras evidências incluem os extensos depósitos de sal marinho e as camadas de pré-sal contendo petróleo derivado de florestas de algas marinhas. A formação destes depósitos requer condições específicas de isolamento de bacias marinhas e rápida evaporação, possivelmente relacionadas a perturbações tectônicas significativas que poderiam ser associadas a eventos de impacto. Da mesma forma, as formações ígneas com pouca sedimentação ou desgaste sobreposto, como o Pão de Açúcar no Rio de Janeiro, sugerem que estes processos geológicos ocorreram em um passado relativamente recente, não tendo havido tempo suficiente para que processos erosivos significativos modificassem suas formas originais.
Trilhões de Pedras Pontiagudas: Evidências de uma Terra Jovem
Um dos argumentos mais intrigantes apresentados em favor de uma cronologia terrestre mais curta é a observação da presença abundante de pedras pontiagudas e angulares na superfície terrestre. Conforme destacado por Sodré (2024), a maioria das rochas expostas mantém bordas afiadas e ângulos pronunciados, características que seriam inconsistentes com longos períodos de exposição a processos erosivos.
Este fenômeno torna-se particularmente significativo quando observamos, em um mesmo terreno, rochas pontiagudas e arredondadas lado a lado, compostas do mesmo material. Se os processos erosivos tiveram tempo suficiente para arredondar completamente algumas rochas, seria esperado que todas as rochas de composição similar no mesmo ambiente fossem igualmente afetadas. A coexistência de pedras angulares e arredondadas sugere, portanto, que as rochas pontiagudas foram expostas à superfície relativamente recentemente, não tendo tido tempo suficiente para sofrer o mesmo grau de erosão que suas contrapartes arredondadas.
A distribuição global destas rochas angulares, ocorrendo em praticamente todos os continentes e em diversos ambientes geológicos, indica que sua origem está relacionada a um ou mais eventos de escala planetária. A hipótese de uma chuva de asteroides recente oferece uma explicação plausível para este fenômeno, sugerindo que o impacto ou impactos múltiplos teriam fragmentado grandes volumes de rocha, produzindo trilhões de fragmentos angulares que não tiveram tempo suficiente desde então para sofrer arredondamento significativo pelos processos normais de intemperismo e erosão.
Rochas Pouco Desgastadas em Áreas de Alta Energia
Outro indicador geológico que sugere uma cronologia terrestre mais recente é a observação de rochas que mostram surpreendentemente pouco desgaste, mesmo quando localizadas em ambientes de alta energia como leitos de rios, bases de cachoeiras e encostas íngremes. Sodré (2017) documentou diversos exemplos deste fenômeno, onde rochas submetidas a condições que deveriam causar erosão rápida e significativa mantêm formas relativamente angulares e superfícies pouco desgastadas.
Em cachoeiras, por exemplo, onde a força abrasiva da água carregada com sedimentos atua continuamente sobre o leito rochoso, seria esperado observar superfícies altamente polidas e estruturas erosivas bem desenvolvidas como marmitas (potholes) se estas formações tivessem sido expostas à ação da água por longos períodos geológicos. No entanto, em muitos casos, as rochas nestas áreas mostram apenas sinais moderados de abrasão, com feições originais ainda claramente discerníveis.
Esta aparente anomalia no grau de erosão observado versus o esperado para longos períodos de tempo representa um desafio significativo para modelos geológicos que postulam idades de milhões ou bilhões de anos para muitas formações rochosas. A hipótese de que estas rochas foram expostas à erosão apenas recentemente, talvez após terem sido deslocadas ou expostas por eventos catastróficos como impactos de asteroides, oferece uma explicação mais consistente com o estado de preservação observado. Esta interpretação alinha-se com a proposta de que grandes partes da geologia terrestre superficial foram significativamente remodeladas por eventos catastróficos em um passado relativamente recente.
Estase Morfológica no Registro Fóssil
Um dos aspectos mais intrigantes do registro fóssil é o fenômeno da estase morfológica – a tendência de muitas espécies de manterem formas essencialmente inalteradas por supostos longos períodos de tempo geológico. De acordo com estudos citados (Simpson, 1944; Benton, 2009), aproximadamente 71% das formas fósseis demonstram esta estase, exibindo pouca ou nenhuma mudança morfológica significativa ao longo de estratos que, segundo a geocronologia convencional, representariam milhões de anos.
Este fenômeno representa um paradoxo quando considerado à luz da observação evolutiva moderna e do conhecimento sobre a influência do ambiente na morfologia dos seres vivos. Em populações atuais, observamos que as características morfológicas respondem relativamente rápido a mudanças ambientais, tanto através de seleção natural quanto por plasticidade fenotípica. Seria esperado, portanto, que ao longo de milhões de anos, com suas inevitáveis flutuações climáticas e ecológicas, as espécies exibissem mudanças morfológicas significativas em resposta a estas pressões ambientais variáveis.
Uma interpretação alternativa para a estase morfológica observada é que ela reflete não uma resistência evolutiva à mudança ao longo de vastos períodos de tempo, mas sim o fato de que os fósseis representam populações que viveram no mesmo período e sob condições ambientais similares. Nesta perspectiva, estratos fósseis supostamente separados por milhões de anos seriam, na realidade, depósitos contemporâneos ou separados por intervalos muito mais curtos, resultantes do sepultamento rápido de ecossistemas inteiros durante eventos catastróficos. Esta interpretação explicaria a preservação de formas idênticas em diferentes estratos sem a necessidade de postular uma improvável resistência à mudança morfológica frente a pressões ambientais variáveis ao longo de milhões de anos.
Implicações da Estase para a Cronologia Terrestre
A estase morfológica generalizada no registro fóssil tem implicações profundas para nossa compreensão da cronologia terrestre. Se 71% das espécies fósseis mostram essencialmente as mesmas formas ao longo de estratos supostamente separados por milhões de anos, como sugerido por Simpson (1944) e confirmado por Benton (2009), então somos confrontados com duas possibilidades interpretativas: ou a evolução morfológica é extraordinariamente resistente a mudanças ambientais durante longos períodos – uma proposição que contradiz observações contemporâneas sobre a plasticidade fenotípica – ou a separação temporal entre os estratos é muito menor do que sugerido pela geocronologia convencional.
A segunda interpretação ganha força quando consideramos o que sabemos sobre a resposta de populações biológicas a mudanças ambientais. Em apenas alguns séculos ou milênios, observamos mudanças morfológicas significativas em populações naturais submetidas a novas pressões seletivas. A ausência dessas mudanças no registro fóssil, portanto, sugere fortemente que os diferentes estratos contendo as mesmas morfologias não representam intervalos de milhões de anos, mas sim deposições geológicas muito mais próximas temporalmente.
Esta reinterpretação do significado da estase morfológica alinha-se com a hipótese de que grandes partes do registro fóssil foram formadas durante eventos catastróficos que sepultaram rapidamente populações inteiras. Sob esta perspectiva, os estratos fossilíferos não representariam uma amostragem esparsa de organismos ao longo de eras geológicas, mas sim “instantâneos” de ecossistemas completos preservados durante eventos de sedimentação rápida, possivelmente associados a impactos de asteroides e seus efeitos secundários, como tsunamis gigantescos e deslizamentos de massa em escala continental.
DNA e Suas Implicações para a Cronologia
Os avanços na análise genética, particularmente nos estudos de degradação do DNA ao longo do tempo, oferecem perspectivas importantes para a discussão sobre a cronologia terrestre. O geneticista John C. Sanford, em colaboração com o geofísico John Baumgardner e outros pesquisadores, tem publicado trabalhos significativos sobre a meia-vida do DNA e suas implicações para o tempo de existência dos organismos na Terra.
O DNA é uma molécula relativamente frágil que sofre degradação espontânea ao longo do tempo, mesmo em condições ideais de preservação. Estudos recentes têm demonstrado que a meia-vida do DNA – o tempo necessário para que metade das ligações em uma amostra se degrade – é relativamente curta em termos geológicos. Mesmo em condições ótimas, como em ossos preservados no permafrost, a meia-vida do DNA foi estimada em apenas algumas centenas a poucos milhares de anos. Esta descoberta impõe limites temporais significativos para a preservação de material genético recuperável.
A implicação direta destas observações é que a recuperação de DNA intacto ou parcialmente intacto de espécimes fósseis, como tem sido relatado em vários estudos recentes, sugere fortemente que estes espécimes não podem ter a antiguidade de milhões de anos atribuída a eles pela geocronologia convencional. A presença de tecidos moles preservados e material genético recuperável em fósseis supostamente antigos representa, portanto, uma evidência significativa favorecendo uma cronologia terrestre substancialmente mais curta, alinhando-se com a hipótese de que eventos como a queda de grandes asteroides ocorreram em um passado muito mais recente do que geralmente se assume.
Aceleração do Decaimento Radioativo por Plasmas
A hipótese de que plasmas de alta energia, gerados durante impactos de asteroides, podem acelerar significativamente as taxas de decaimento radioativo representa um desafio fundamental para os pressupostos da geocronologia radiométrica convencional. Este mecanismo oferece uma explicação plausível para discrepâncias observadas entre diferentes métodos de datação e para anomalias geocronológicas documentadas em diversas regiões.
O princípio fundamental dos métodos de datação radiométrica é a constância das taxas de decaimento radioativo em sistemas fechados. No entanto, pesquisas em física nuclear têm demonstrado que fatores externos podem, sob certas condições, influenciar significativamente estas taxas. Técnicas patenteadas de descontaminação nuclear, por exemplo, utilizam aceleradores de partículas para induzir o decaimento acelerado de isótopos radioativos, demonstrando inequivocamente que o decaimento radioativo não é absolutamente imutável como frequentemente se assume.
Os plasmas gerados durante impactos de asteroides representam ambientes físicos extremos, caracterizados por temperaturas que podem exceder milhões de graus Celsius, intensos campos eletromagnéticos e fluxos de partículas de alta energia. Nestas condições, os princípios convencionais da física nuclear podem ser temporariamente alterados. Elétrons acelerados a velocidades relativísticas podem superar a barreira de Coulomb, facilitando interações nucleares que normalmente seriam energeticamente desfavoráveis. Além disso, as altas temperaturas e pressões podem aumentar significativamente a probabilidade de tunelamento quântico através da barreira de Coulomb, potencialmente aumentando as taxas de decaimento por várias ordens de magnitude.
Implicações para a Geocronologia Radiométrica
As implicações da possível aceleração do decaimento radioativo durante eventos de impacto são profundas para a geocronologia radiométrica. Se os plasmas gerados por impactos de asteroides podem temporariamente aumentar as taxas de decaimento, então rochas expostas a estes plasmas experimentariam um “envelhecimento radiométrico acelerado” – desenvolvendo proporções isotópicas que sugeririam idades muito maiores do que seu tempo real de existência quando analisadas por métodos convencionais.
Este fenômeno poderia explicar várias anomalias observadas em datações radiométricas, como discrepâncias sistemáticas entre diferentes métodos aplicados à mesma amostra, ou resultados inconsistentes com evidências geológicas, paleontológicas ou arqueológicas. Por exemplo, a datação de rochas vulcânicas associadas a artefatos humanos conhecidos frequentemente produz idades radiométricas muitas vezes maiores do que a idade historicamente documentada dos artefatos, um fenômeno que poderia ser explicado por episódios de decaimento acelerado no passado.
A hipótese do decaimento acelerado também oferece uma perspectiva alternativa sobre o significado das “idades absolutas” derivadas de métodos radiométricos. Em vez de representarem o tempo real decorrido desde a formação da rocha, estas idades poderiam refletir uma combinação do tempo cronológico e dos efeitos de episódios de decaimento acelerado. Esta reinterpretação sugere que a escala de tempo geológica convencional pode estar significativamente expandida em relação à cronologia real da Terra, com implicações profundas para nossa compreensão da história planetária e da evolução da vida.
Coincidência Temporal de Eventos Geologicamente Distintos
Um aspecto particularmente intrigante que questiona a cronologia geológica convencional é a observação de coincidências temporais entre eventos que, teoricamente, deveriam estar separados por milhões de anos. Um exemplo notável desta coincidência é a relação entre o impacto de Chicxulub, na península de Yucatán, e as erupções do Deccan Traps na Índia.
Richards (2015) apresentou evidências convincentes de que o impacto de Chicxulub, tradicionalmente associado à extinção do Cretáceo-Paleogeno (K-Pg), pode ter desencadeado ou intensificado significativamente a atividade vulcânica que produziu os enormes derrames basálticos do Deccan. Esta correlação temporal entre um impacto de asteroide e um episódio de vulcanismo de grande escala desafia a noção convencional de que estes eventos representariam processos geológicos independentes, separados por milhões de anos.
A implicação desta observação é profunda: se eventos que a geocronologia convencional coloca em diferentes períodos geológicos mostram evidências de correlação causal direta, então a escala de tempo atribuída a estes eventos deve ser significativamente comprimida. Esta compressão temporal alinha-se com a hipótese de que grande parte da história geológica da Terra, como registrada em suas rochas e fósseis, representa não uma progressão lenta ao longo de bilhões de anos, mas sim uma sequência de eventos catastróficos ocorridos em um passado relativamente recente. A impossibilidade lógica de que eventos causalmente relacionados estejam separados por milhões de anos fornece um argumento consistente para a revisão das escalas de tempo geológicas convencionais.
Preservação de Tecidos Moles em Fósseis “Antigos”
A descoberta de tecidos moles preservados em fósseis supostamente antigos representa um dos desafios mais significativos à geocronologia convencional. Exemplos notáveis incluem os achados de Mary Schweitzer e colaboradores, que identificaram tecidos moles com elasticidade, vasos sanguíneos, células e até mesmo proteínas preservadas em ossos de dinossauros como Tyrannosaurus rex, convencionalmente datados em 68 milhões de anos.
Armitage (2013) destacou este paradoxo em seu estudo sobre tecidos moles preservados em um chifre de Triceratops horridus. Como ele apropriadamente questionou: “O que também não está claro é como essas estruturas de biofilme poderiam sobreviver à deterioração do tempo, uma vez que outros microrganismos poderiam começar a digerir inclusive esses”. Esta observação aponta para a dificuldade fundamental em reconciliar a preservação de materiais biológicos altamente degradáveis com as idades de dezenas de milhões de anos atribuídas a estes fósseis.
O conhecimento atual sobre a degradação de biomoléculas indica que proteínas, lípidos e outras macromoléculas degradam-se em escalas de tempo de milhares, não milhões, de anos, mesmo em condições ideais de preservação. A presença de tecidos ainda flexíveis, com estruturas celulares reconhecíveis e biomoléculas originais preservadas, é praticamente inexplicável dentro do paradigma temporal convencional. Estes achados sugerem fortemente que os fósseis em questão são muito mais recentes do que as idades radiométricas indicam, corroborando a hipótese de que eventos como a extinção dos dinossauros ocorreram em um passado geológico muito mais recente, possivelmente relacionado a impactos de asteroides que também aceleraram o decaimento radioativo das rochas circundantes.
Incongruências nas Datações Radiométricas
As incongruências nas datações radiométricas de rochas adjacentes ou geologicamente relacionadas representam outro indicador significativo que questiona a confiabilidade dos métodos de datação absoluta. Numerosos casos têm sido documentados onde rochas que, por sua relação estratigráfica ou contexto geológico, deveriam ter idades similares, mostram idades radiométricas drasticamente diferentes quando analisadas pelos mesmos métodos.
Estas discrepâncias são particularmente evidentes em contextos vulcânicos, onde fluxos de lava que comprovadamente se formaram durante a mesma erupção frequentemente produzem idades radiométricas que variam em milhares ou até milhões de anos. De forma similar, diferentes métodos radiométricos aplicados à mesma amostra frequentemente geram resultados inconsistentes, um fenômeno que levou ao desenvolvimento de técnicas de “filtragem” de dados que selecionam apenas os resultados considerados “geologicamente plausíveis” – uma prática que introduz circularidade no processo de datação.
Estas incongruências são difíceis de explicar dentro do paradigma convencional que assume taxas de decaimento constantes em sistemas fechados. No entanto, elas se tornam compreensíveis quando consideramos a possibilidade de que diferentes rochas possam ter sido expostas a diferentes intensidades de plasmas e campos eletromagnéticos durante eventos de impacto, resultando em graus variáveis de aceleração do decaimento radioativo. Esta hipótese oferece uma explicação coerente para as variações aparentemente aleatórias em idades radiométricas observadas em rochas geologicamente relacionadas, sugerindo que estas idades refletem não o tempo real de formação, mas sim os efeitos diferenciais de processos de aceleração do decaimento em diferentes materiais e localizações.
Afunilamento Populacional em Análises Genéticas
Um aspecto particularmente revelador do registro genético humano é o evidente afunilamento populacional que estudos genéticos têm consistentemente identificado. Chang (2004) e outros pesquisadores demonstraram, através de análises estatísticas do DNA humano atual, que nossa espécie experimentou uma redução drástica em sua diversidade genética há aproximadamente 4.000 anos, indicando uma população efetiva extremamente reduzida neste período.
Este fenômeno de afunilamento genético (bottleneck) é evidenciado pela surpreendente homogeneidade genética da humanidade moderna quando comparada com outras espécies de mamíferos. Apesar de nossa população global atual exceder 7 bilhões de indivíduos, nossa diversidade genética é comparável à de espécies ameaçadas com populações muito menores – um paradoxo que só pode ser explicado por um evento catastrófico recente que reduziu drasticamente a população humana a um pequeno número de indivíduos reprodutores.
As implicações deste afunilamento populacional são significativas para a discussão sobre catástrofes geológicas recentes. A coincidência temporal entre o afunilamento genético humano e evidências de grandes impactos de asteroides sugere uma possível relação causal: um evento catastrófico global, como uma chuva de asteroides, poderia ter dizimado a população humana, deixando apenas pequenos grupos isolados de sobreviventes. Esta hipótese é corroborada pela observação de que grupos étnicos geograficamente próximos frequentemente mostram notável semelhança fenotípica, sugerindo origem a partir de populações fundadoras muito pequenas e relacionadas, exatamente o padrão esperado após uma catástrofe global seguida por recolonização a partir de grupos de sobreviventes isolados.
Biossemelhança Étnica e Evidências de Catástrofe
A notável semelhança fenotípica observada entre indivíduos pertencentes a grupos étnicos geograficamente próximos fornece evidências adicionais de um evento catastrófico recente que reduziu drasticamente a população humana. Grupos como os diversos povos de origem africana, ou as populações mongoloides do leste asiático (chineses, japoneses e coreanos), exibem graus de similaridade física que sugerem derivação a partir de populações fundadoras extremamente pequenas e geneticamente relacionadas.
Este padrão de biossemelhança étnica é difícil de explicar dentro de um modelo de evolução humana gradual ao longo de centenas de milhares de anos. Se as populações humanas tivessem mantido tamanhos significativos por longos períodos, esperaríamos observar muito maior diversidade fenotípica dentro de cada grupo étnico. A relativa homogeneidade observada é mais consistente com um cenário onde pequenos grupos de sobreviventes, após um evento catastrófico global, reproduziram-se em isolamento relativo, levando a uma rápida fixação de características fenotípicas específicas através de efeitos fundadores e deriva genética em populações pequenas.
Intrigantemente, descobertas arqueológicas e paleontológicas em várias regiões do mundo têm revelado padrões de distribuição étnica histórica diferentes dos atuais. Por exemplo, Lund (1801-1880) documentou fósseis com padrão morfológico negróide na América do Sul, uma região onde as populações indígenas pré-colonização européia eram predominantemente de características mongoloides. Achados semelhantes foram reportados no México e em São Paulo. Da mesma forma, na Ásia, existem evidências de populações antigas com características negróides e caucasianas em áreas atualmente dominadas por povos mongoloides. Este padrão de substituição populacional completa, sem evidências de miscigenação significativa, é consistente com um cenário de extinção catastrófica das populações originais, seguida por recolonização por grupos sobreviventes de outras regiões – exatamente o que seria esperado após um evento catastrófico de escala global.
Endogamia e Sobrevivência Populacional Pós-Catástrofe
O cenário de uma catástrofe global recente que reduziu drasticamente a população humana levanta questões importantes sobre como pequenos grupos isolados de sobreviventes conseguiram superar os desafios genéticos associados à endogamia. Em populações extremamente reduzidas, a reprodução entre indivíduos geneticamente relacionados tipicamente resulta em depressão endogâmica – a expressão aumentada de alelos deletérios recessivos, levando à redução da aptidão biológica e potencialmente à extinção.
Uma possível explicação para a sobrevivência de pequenas populações fundadoras após um evento catastrófico envolve a consideração da entropia genética e taxas de mutação. Se as populações humanas pré-catástrofe tivessem acumulado relativamente poucas mutações deletérias – consistente com as evidências de baixas taxas de mutação em múmias antigas comparadas à população atual – então o risco de depressão endogâmica seria significativamente reduzido. Populações com genomas relativamente “limpos” de alelos deletérios seriam muito mais capazes de sobreviver a extremos afunilamentos populacionais sem sucumbir aos efeitos negativos da endogamia.
Esta hipótese harmoniza-se com observações sobre o relativamente baixo número de mutações encontradas em DNA antigo quando comparado com populações modernas. A análise de DNA mitocondrial de múmias e outros restos humanos antigos revela aproximadamente 300 mutações mitocondriais, em contraste com as cerca de 20.000 mutações catalogadas no banco de dados MITOMAP para humanos contemporâneos. Esta disparidade sugere que o evento catastrófico não apenas reduziu a população humana, mas também pode ter exposto os sobreviventes a níveis elevados de radiação que aceleraram significativamente as taxas de mutação, explicando o grande aumento observado na carga mutacional entre populações antigas e modernas.
Disparidade no Acúmulo de Mutações ao Longo da História
Um dos aspectos mais intrigantes revelados por estudos genéticos recentes é a notável disparidade no número de mutações observadas entre amostras de DNA antigo e populações humanas contemporâneas. Esta disparidade representa um desafio significativo para modelos convencionais de acúmulo gradual e constante de mutações ao longo do tempo evolutivo.
Análises de DNA extraído de múmias egípcias e outros restos humanos antigos, com idade aproximada de 5.000 anos segundo cronologias arqueológicas convencionais, revelam aproximadamente 300 mutações mitocondriais (Mmt). Em contraste, o banco de dados MITOMAP, que cataloga mutações no DNA mitocondrial humano atual, já documentou cerca de 20.000 Mmt na população mundial contemporânea. Esta disparidade de quase 70 vezes no número de mutações é difícil de reconciliar com as taxas conhecidas de acúmulo de mutações por geração, estimadas em aproximadamente 0,024 Mmt por geração.
Baseando-se nestas taxas de mutação observadas empiricamente, seriam necessárias dezenas ou centenas de milhares de gerações para explicar a diferença entre o número de mutações em amostras antigas e modernas – um intervalo temporal claramente incompatível com os aproximadamente 200-250 gerações que separam as múmias de 5.000 anos da população atual. Esta incongruência sugere fortemente que um ou mais eventos no passado relativamente recente causaram um aumento drástico e temporário na taxa de mutação. A exposição a níveis elevados de radiação ionizante durante e após impactos de asteroides representa uma explicação plausível para este padrão, consistente com a hipótese de que estes eventos ocorreram em um passado muito mais recente do que geralmente se assume.
Taxas de Mutação e Consequências para a Cronologia
As implicações das discrepâncias observadas nas taxas de acúmulo de mutações são profundas para nossa compreensão da cronologia humana e, por extensão, da história terrestre recente. Se considerarmos as taxas empiricamente estabelecidas de acúmulo de mutações mitocondriais (aproximadamente 0,024 Mmt por geração) e aplicarmos estas taxas à diferença observada entre amostras antigas (300 Mmt) e contemporâneas (20.000 Mmt), chegamos a uma conclusão surpreendente.
Para acumular aproximadamente 19.700 mutações adicionais às taxas normais, seriam necessárias cerca de 820.833 gerações (19.700 ÷ 0,024). Assumindo uma duração média de geração humana de 25 anos, isto corresponde a aproximadamente 20,5 milhões de anos – um período claramente incompatível com a existência conhecida de nossa espécie e com a idade estimada das múmias analisadas. Esta discrepância monumental sugere que o acúmulo de mutações não ocorreu gradualmente ao longo do tempo, mas foi dramaticamente acelerado por um ou mais eventos que aumentaram temporariamente as taxas de mutação por ordens de magnitude.
A exposição a níveis elevados de radiação ionizante, como aquela que ocorreria durante e após impactos significativos de asteroides, representa um mecanismo plausível para este aumento dramático nas taxas de mutação. A radiação ionizante é um conhecido agente mutagênico que causa danos diretos ao DNA, resultando em mutações pontuais, deleções, inserções e rearranjos cromossômicos. Se a população humana foi exposta a níveis significativamente elevados de radiação há alguns milhares de anos, como resultado de impactos de asteroides, isto explicaria o padrão observado de acúmulo desproporcional de mutações em um período relativamente curto.
Predominância de SNPs nas Mutações Humanas
Um aspecto particularmente revelador do padrão de mutações observado na população humana contemporânea é a predominância de polimorfismos de nucleotídeo único (SNPs – Single Nucleotide Polymorphisms). Estes SNPs representam a forma mais comum de variação genética entre indivíduos, caracterizando-se pela alteração de uma única base nitrogenada na sequência de DNA.
A prevalência de SNPs em relação a outros tipos de mutações, como deleções, inserções ou inversões de segmentos maiores, é consistente com os efeitos conhecidos da radiação ionizante no material genético. Quando células são expostas à radiação, particularmente raios gama e raios X, os danos mais frequentes ao DNA são quebras de fita simples que, quando reparadas incorretamente, frequentemente resultam em substituições de nucleotídeos únicos – precisamente o padrão observado nos SNPs.
Esta concordância entre o tipo predominante de mutações na população humana e o padrão característico de danos ao DNA induzidos por radiação fornece evidência adicional apoiando a hipótese de exposição a níveis elevados de radiação no passado recente. Se a população humana foi submetida a um ambiente altamente radioativo, como aquele que seguiria impactos significativos de asteroides, esperaríamos observar exatamente o padrão que de fato encontramos: um número anormalmente elevado de mutações, com predominância de SNPs sobre outros tipos de alterações genéticas. Esta observação corrobora a proposta de que grandes impactos de asteroides ocorreram em um passado relativamente recente, expondo a população humana sobrevivente a níveis de radiação suficientes para causar um aumento dramático no número de mutações em um período relativamente curto.
Separação Continental e Endemismo
A distribuição biogeográfica de espécies endêmicas fornece evidências significativas que podem ser interpretadas como suporte para a hipótese de catástrofes geológicas recentes. O alto grau de endemismo observado em diferentes continentes – a ocorrência de espécies exclusivas a determinadas regiões geográficas – sugere que a separação dos continentes pode ter ocorrido mais rapidamente do que proposto pelos modelos convencionais de deriva continental.
De acordo com a hipótese apresentada, os plasmas de alta energia gerados por impactos de asteroides podem ter contribuído para fissurar a crosta continental da Terra, iniciando um processo de separação continental tão rápido que não houve tempo suficiente para a distribuição homogênea dos seres vivos. Esta aceleração do processo de deriva continental, combinada com a extinção catastrófica de grande parte da biota terrestre, resultaria precisamente no padrão biogeográfico que observamos hoje: continentes separados com biotas distintas, caracterizadas por alto grau de endemismo.
Um exemplo notável que apoia esta interpretação é a fauna australiana, onde aproximadamente 80% das espécies são endêmicas. Este padrão é difícil de explicar através de modelos graduais de especiação ao longo de milhões de anos, especialmente considerando que a Austrália se separou de outros continentes há supostamente 40-50 milhões de anos. Se este intervalo de tempo fosse preciso, esperaríamos observar muito maior convergência evolutiva e intercâmbio de espécies. O alto grau de endemismo sugere, ao contrário, que a separação continental ocorreu recentemente o suficiente para que as populações isoladas não tivessem tempo de divergir significativamente além do nível de espécie ou gênero, mantendo semelhanças taxonômicas em níveis superiores (família, ordem) que refletem ancestralidade comum recente.
Casos Notáveis de Endemismo Continental
Inúmeros exemplos de endemismo continental fornecem evidências adicionais que sugerem uma separação geológica relativamente recente dos continentes. Além do caso já mencionado da fauna australiana, podemos observar padrões similares em outros continentes e ilhas isoladas, onde espécies ou grupos de espécies desenvolveram-se exclusivamente em determinadas áreas geográficas.
Na América do Sul, plantas como a jabuticabeira (Plinia cauliflora) representam exemplos notáveis de endemismo. Esta espécie, nativa das florestas do Brasil, desenvolveu características únicas que não são encontradas em nenhum outro lugar do mundo. Da mesma forma, a fauna sul-americana inclui grupos inteiros que evoluíram isoladamente, como os marsupiais didelfídeos (gambás americanos) e os primatas do Novo Mundo (Platyrrhini), que divergiram significativamente de seus parentes em outros continentes.
Em Madagascar, uma ilha que se separou do continente africano há supostamente 165 milhões de anos, observamos um grau extraordinário de endemismo, com mais de 90% de sua fauna sendo exclusiva à ilha. Este isolamento extremo resultou no desenvolvimento de grupos únicos como os lêmures, que não são encontrados naturalmente em nenhum outro lugar do planeta. O mesmo padrão se repete nas Ilhas Galápagos, Havaí, e outras ilhas oceânicas isoladas, onde processos de especiação endêmica produziram biotas altamente distintivas. Estes exemplos de isolamento biogeográfico e subsequente endemismo são consistentes com a hipótese de uma separação continental relativamente recente, onde as populações ancestrais sobreviventes a uma catástrofe global não tiveram tempo suficiente para se dispersar amplamente antes que barreiras geográficas significativas se estabelecessem.
Hipótese de Chuva de Asteroides Recente
A convergência de múltiplas linhas de evidência independentes – desde anomalias na geocronologia radiométrica até padrões biogeográficos e genéticos – sugere fortemente a ocorrência de um evento catastrófico global em um passado relativamente recente. A hipótese de uma chuva de asteroides ocorrida há aproximadamente 4.000-5.000 anos oferece um mecanismo unificador que potencialmente explica esta diversidade de observações aparentemente díspares.
De acordo com esta hipótese, a Terra foi submetida a múltiplos impactos de asteroides em um período relativamente curto, possivelmente resultantes da fragmentação de um corpo celeste maior ou da passagem do sistema solar por uma região do espaço com maior densidade de detritos cósmicos. Estes impactos teriam produzido efeitos devastadores em escala global: ondas de choque, tsunamis gigantescos, incêndios generalizados, escurecimento atmosférico, mudanças climáticas abruptas e, criticamente, intensos plasmas de alta energia que poderiam ter acelerado processos de decaimento radioativo e gerado níveis elevados de radiação ionizante.
Os efeitos geológicos destes impactos incluiriam a formação rápida de extensas camadas sedimentares, a criação de numerosas crateras de impacto (muitas das quais já erodidas ou obscurecidas), possível aceleração da deriva continental através da fragmentação da crosta terrestre, e o “envelhecimento” aparente de rochas expostas aos plasmas de impacto. Os efeitos biológicos seriam igualmente profundos: extinção massiva de espécies, redução drástica das populações sobreviventes a pequenos grupos isolados, e exposição dos organismos remanescentes a níveis elevados de radiação, resultando em aumento significativo nas taxas de mutação e subsequente diversificação genética acelerada a partir de populações fundadoras reduzidas.
Evidências de Impactos Múltiplos
A hipótese de uma chuva de asteroides recente, em oposição a um impacto isolado, é suportada por evidências de múltiplos eventos de impacto aproximadamente contemporâneos em diferentes regiões do planeta. Estudos detalhados de crateras de impacto conhecidas revelam clusters temporais que sugerem eventos relacionados, possivelmente resultantes da fragmentação de um corpo celeste maior antes de atingir a Terra.
Ormö et al. (2014) documentaram evidências do primeiro impacto conhecido de um asteroide binário na Terra, demonstrando que impactos múltiplos e relacionados podem ocorrer simultaneamente. Este tipo de evento, multiplicado em escala, poderia produzir precisamente o padrão de impactos distribuídos globalmente que a hipótese propõe. Além disso, Hassler e Simonson (2001) analisaram registros sedimentares de impactos extraterrestres, identificando “horizontes de impacto” que contêm evidências de múltiplos eventos aproximadamente contemporâneos.
Glikson et al. (2004) forneceram evidências adicionais ao identificar múltiplas unidades de fallout de impactos antigos no Cráton de Pilbara, na Austrália Ocidental. Estas camadas de ejecta, tradicionalmente interpretadas como separadas por centenas de milhões de anos, apresentam semelhanças composicionais e estruturais que sugerem possível contemporaneidade quando reinterpretadas à luz da hipótese de aceleração do decaimento radioativo. Similarmente, Heck et al. (2017) investigaram meteoritos raros comuns no período Ordoviciano, identificando evidências de um aumento significativo no fluxo de material extraterrestre durante este período, consistente com a ocorrência de uma chuva de asteroides ou fragmentação de um corpo parental maior.
Proposta de Pesquisa para Validação da Hipótese
Para avançar além do estágio de hipótese e desenvolver um modelo científico robusto e testável, propõe-se uma abordagem multidisciplinar focada em três linhas principais de investigação que poderiam fornecer evidências confirmatórias ou refutatórias para a hipótese da chuva de asteroides recente.
Primeiramente, sugere-se uma análise sistemática dos ângulos de impacto em crateras conhecidas, tanto na Terra quanto na Lua. Se múltiplos asteroides atingiram o sistema Terra-Lua em um período relativamente curto, como parte de um enxame de fragmentos relacionados, esperaríamos observar padrões não aleatórios nos ângulos de impacto. Especificamente, crateras formadas por fragmentos de um mesmo corpo parental tenderiam a mostrar trajetórias de impacto consistentes com uma origem comum no espaço. Técnicas modernas de análise morfométrica de crateras, combinadas com modelagem computacional, permitem estimar com razoável precisão os ângulos e direções de impacto a partir da geometria das crateras e da distribuição de ejecta.
Em segundo lugar, propõe-se a caracterização detalhada da composição química e isotópica de materiais de impacto de diferentes crateras, visando identificar assinaturas que indiquem origem a partir de um mesmo corpo parental. Asteroides de uma mesma família compartilham características composicionais distintivas que podem ser detectadas em seus fragmentos terrestres. Por exemplo, a cratera de Chicxulub tem sido associada à família de asteroides Baptistina por alguns pesquisadores, baseando-se em similaridades composicionais. A identificação de assinaturas químicas e isotópicas compartilhadas entre materiais de diferentes crateras forneceria evidência forte para sua origem comum.
Previsões Testáveis da Hipótese
Uma característica fundamental de qualquer hipótese científica robusta é sua capacidade de gerar previsões testáveis que possam ser confirmadas ou refutadas por observações e experimentos. A hipótese da chuva de asteroides recente oferece várias previsões específicas que podem ser investigadas por pesquisas futuras.
Uma previsão crítica relaciona-se à terceira linha de investigação sugerida: se múltiplos asteroides impactaram a Terra como parte de um enxame de fragmentos relacionados há alguns milhares de anos, então outros fragmentos do mesmo enxame que não colidiram com a Terra ou a Lua devem ainda estar presentes no sistema solar interior. Baseando-se em análises de ângulos de impacto e composição de materiais, seria possível calcular retrospectivamente as órbitas originais destes fragmentos e, por extensão, prever as órbitas atuais dos fragmentos sobreviventes. A identificação de uma família de asteroides ou cometas com órbitas consistentes com estas previsões e composição similar aos materiais de impacto forneceria evidência substancial para a hipótese.
Outra previsão testável relaciona-se aos efeitos da radiação proposta nos organismos sobreviventes. Se a hipótese estiver correta, deveríamos observar assinaturas genéticas de exposição à radiação em populações descendentes de sobreviventes do evento. Especificamente, esperaríamos encontrar evidências de “seleção de purificação” intensa atuando sobre genes relacionados a mecanismos de reparo de DNA e resistência à radiação nas gerações imediatamente posteriores ao evento, seguidas por relaxamento desta pressão seletiva em gerações mais recentes. Estudos de genômica populacional comparativa poderiam potencialmente detectar estes sinais de seleção natural associados a resistência à radiação.
Sinais Texturais e Sedimentológicos de Catástrofes
A superfície terrestre está repleta de evidências sedimentológicas e texturais que sugerem a ocorrência de eventos catastróficos de grande magnitude em um passado geológico relativamente recente. Estas características incluem megamarcas onduladas (megaripples), camadas de sedimentos caóticos, depósitos de megainundações e outras formações que dificilmente podem ser explicadas por processos graduais operando em escalas de tempo convencionais.
Berthault, Lalomov e Tugarova (2011) realizaram estudos detalhados de reconstrução das condições paleohidrodinâmicas de formação de arenitos Cambro-Ordovicianos na plataforma Russa Noroeste. Suas análises revelaram evidências de deposição rápida em condições de alta energia, incompatíveis com a interpretação convencional de acumulação lenta ao longo de milhões de anos. Características como estratificação cruzada em escala gigante, ausência de bioturbação em interfaces entre camadas, e preservação de grãos angulares em arenitos supostamente antigos sugerem deposição catastrófica seguida por litificação rápida, em vez de processos graduais de sedimentação e diagênese.
Estudos similares em outras formações sedimentares, desde o Ediacarano-Cambriano até o Pleistoceno, revelam padrões consistentes com a hipótese de eventos catastróficos recentes. A presença de camadas sedimentares extensas e contínuas, frequentemente estendendo-se por centenas ou milhares de quilômetros com espessura e características texturais relativamente uniformes, sugere deposição durante eventos de alta energia capazes de transportar e distribuir sedimentos em escala continental. Tais eventos poderiam incluir megainundações, tsunamis continentais e outras perturbações hidrodinâmicas massivas potencialmente desencadeadas por impactos de asteroides.
Mares de Sal e Camadas de Pré-Sal
Os extensos depósitos de sal marinho e as formações de pré-sal contendo reservas significativas de petróleo representam enigmas geológicos cuja formação pode ser mais adequadamente explicada no contexto de eventos catastróficos de escala global. Estas formações, tradicionalmente interpretadas como resultado de processos lentos ocorrendo ao longo de milhões de anos, apresentam características que sugerem episódios de rápida deposição e condições ambientais extremas.
Os depósitos de sal marinho, como aqueles encontrados sob o Mar Mediterrâneo (Formação Messinian) ou no Golfo do México, requerem a evaporação de volumes extraordinários de água salgada. Para formar camadas de sal com espessuras de centenas de metros, seria necessário evaporar bacias marinhas inteiras repetidamente. No paradigma convencional, este processo é atribuído a ciclos de isolamento e reinundação de bacias ao longo de períodos extensos. No entanto, a homogeneidade composicional destes depósitos e a ausência de camadas significativas de materiais terrígenos intercalados sugerem deposição contínua e rápida, mais consistente com evaporação catastrófica possivelmente relacionada a aquecimento atmosférico extremo seguindo impactos de asteroides.
Da mesma forma, as camadas de pré-sal, famosas por seus reservatórios de petróleo ao longo da costa brasileira e africana, contêm evidências de deposição rápida de material orgânico derivado principalmente de algas marinhas. A preservação em larga escala deste material orgânico, sua conversão em hidrocarbonetos, e seu sepultamento sob espessas camadas de sal sugerem uma sequência de eventos catastróficos: mortalidade em massa de organismos marinhos, possivelmente devido a condições ambientais extremas, seguida por rápido sepultamento e subsequente formação de camadas de sal. Este cenário é consistente com as perturbações ambientais globais que seguiriam múltiplos impactos de asteroides.
Formações Ígneas e Intemperismo Limitado
As formações ígneas expostas em várias partes do mundo, muitas das quais mostram sinais surpreendentemente limitados de intemperismo e erosão, fornecem evidências adicionais que sugerem uma cronologia terrestre mais curta do que convencionalmente aceito. Exemplos notáveis incluem estruturas massivas como o Pão de Açúcar no Rio de Janeiro e outras formações graníticas e basálticas que mantêm superfícies relativamente frescas e formas angulares, apesar de supostamente terem sido expostas a processos intempéricos por milhões ou até bilhões de anos.
Se estas formações realmente tivessem idades na ordem de centenas de milhões de anos, esperaríamos observar graus muito mais avançados de decomposição química e desintegração física. Granitos e outras rochas ígneas expostas a condições atmosféricas normais por longos períodos tipicamente desenvolvem crostas de intemperismo profundas, com minerais primários como feldspatos e micas sendo convertidos em argilominerais. No entanto, muitas destas formações mostram apenas camadas superficiais de alteração, com rocha fresca e inalterada a poucos centímetros da superfície.
A presença de estruturas ígneas com intemperismo limitado, frequentemente em proximidade a depósitos sedimentares que, segundo a interpretação convencional, teriam se acumulado após a formação das rochas ígneas, representa uma inconsistência cronológica significativa. Esta observação é mais facilmente explicada assumindo que tanto as formações ígneas quanto os depósitos sedimentares são relativamente recentes, possivelmente resultantes ou expostos durante eventos catastróficos como a proposta chuva de asteroides. Neste cenário, o intemperismo limitado refletiria simplesmente o intervalo de tempo relativamente curto desde a exposição destas rochas à superfície.
Relevância da Hipótese para a Ciência Contemporânea
A hipótese de impactos de asteroides recentes como fator significativo na geologia terrestre, geocronologia e evolução biológica tem implicações profundas para diversas disciplinas científicas. Longe de ser meramente uma proposta alternativa isolada, esta hipótese intersecta campos como astrofísica, geologia, paleontologia, genética e arqueologia, oferecendo potencial para sínteses interdisciplinares inovadoras e resolução de anomalias persistentes em cada uma destas áreas.
No campo da geologia, a consideração dos efeitos de plasmas de impacto na aceleração do decaimento radioativo poderia resolver discrepâncias bem documentadas entre diferentes métodos de datação radiométrica aplicados às mesmas amostras. Similarmente, a hipótese oferece uma explicação plausível para a preservação anômala de tecidos moles e biomoléculas em fósseis supostamente antigos, um fenômeno que continua a desafiar explicações convencionais. Na genética evolutiva, o conceito de um evento recente causando um aumento dramático nas taxas de mutação poderia explicar a discrepância entre o número de mutações observadas em amostras antigas versus contemporâneas.
Além de seu potencial explanatório, a hipótese gera previsões testáveis que podem ser investigadas usando tecnologias e metodologias científicas contemporâneas. Avanços em técnicas analíticas de alta precisão, simulações computacionais sofisticadas e capacidades ampliadas de observação astronômica permitem testar aspectos específicos da hipótese que seriam inacessíveis a gerações anteriores de cientistas. Esta combinação de relevância interdisciplinar e testabilidade empírica posiciona a hipótese como uma contribuição potencialmente valiosa ao discurso científico contemporâneo, merecedora de investigação rigorosa independentemente de sua divergência de paradigmas estabelecidos.
Desafios à Hipótese e Possíveis Refutações
Como qualquer proposta científica, a hipótese de impactos de asteroides recentes e seus efeitos na geocronologia e evolução biológica enfrenta desafios significativos e potenciais refutações que devem ser considerados criticamente. O reconhecimento destes desafios é essencial para o desenvolvimento de uma abordagem verdadeiramente científica, baseada em evidências e aberta à falsificação.
Um desafio fundamental relaciona-se à universalidade da aceleração do decaimento radioativo proposta. Se impactos de asteroides realmente causaram aceleração significativa do decaimento, esperaríamos observar padrões consistentes de “envelhecimento” aparente em rochas afetadas. No entanto, a literatura geocronológica documenta numerosos casos onde diferentes métodos radiométricos (urânio-chumbo, potássio-argônio, rubídio-estrôncio, etc.) aplicados à mesma amostra produzem resultados concordantes. Esta concordância entre métodos baseados em diferentes sistemas isotópicos seria altamente improvável se a aceleração do decaimento afetasse todos os isótopos indiscriminadamente.
Outro desafio significativo vem de registros astronômicos e geológicos de longo prazo. Calendários astronômicos antigos, registros de eclipses, e varves (camadas sedimentares anuais) em lagos glaciais fornecem evidências de continuidade cronológica que seriam difíceis de reconciliar com um evento catastrófico global ocorrido há apenas alguns milhares de anos. Além disso, a existência de sistemas ecológicos altamente especializados e interdependentes, como recifes de coral com histórias evolutivas aparentemente longas, representa um desafio para a hipótese de perturbação global recente. Qualquer desenvolvimento futuro da hipótese precisará abordar estes desafios, propondo mecanismos específicos pelos quais certos registros cronológicos poderiam ser preservados mesmo durante eventos catastróficos de larga escala.
Papel da Ciência Interdisciplinar na Avaliação da Hipótese
A avaliação adequada da hipótese de impactos de asteroides recentes e seus efeitos na geologia terrestre, geocronologia e evolução biológica exige uma abordagem genuinamente interdisciplinar que transcenda as fronteiras tradicionais entre campos científicos. A natureza da hipótese, que atravessa domínios desde a astrofísica até a genética molecular, requer colaboração entre especialistas de diversas áreas para desenvolver metodologias de teste apropriadas e interpretar resultados com competência.
A colaboração entre geofísicos, geoquímicos e físicos nucleares é particularmente crucial para investigar a possibilidade de aceleração do decaimento radioativo em condições de plasma de alta energia. Experimentos laboratoriais simulando condições extremas semelhantes àquelas geradas durante impactos, combinados com modelagem teórica detalhada dos processos nucleares envolvidos, poderiam fornecer insights valiosos sobre a plausibilidade física deste mecanismo proposto. Similarmente, a cooperação entre paleontólogos, geneticistas e especialistas em biomoléculas é essencial para avaliar a significância da preservação de tecidos moles e DNA em fósseis antigos.
A integração de dados de múltiplas disciplinas também permite a triangulação de evidências, fortalecendo ou enfraquecendo a hipótese através de convergência ou divergência de resultados independentes. Por exemplo, se análises astronômicas de órbitas de asteroides contemporâneos identificarem uma família consistente com origem comum há alguns milhares de anos, e simultaneamente estudos geológicos detalhados de crateras terrestres revelarem padrões consistentes com impactos múltiplos da mesma época, e ainda análises genéticas identificarem sinais de radiação elevada no mesmo período – tal convergência de evidências independentes fortaleceria significativamente a hipótese. Inversamente, contradições fundamentais entre evidências de diferentes campos poderiam indicar necessidade de revisão ou abandono da hipótese.
Implicações para Pesquisas Futuras em Geocronologia
Independentemente da validade última da hipótese específica de impactos de asteroides recentes, sua formulação levanta questões importantes sobre os fundamentos da geocronologia moderna e sugere direções produtivas para pesquisas futuras neste campo crítico para as ciências da Terra.
Uma implicação significativa é a necessidade de investigação mais aprofundada sobre a estabilidade das taxas de decaimento radioativo sob condições extremas. Embora a física nuclear convencional considere estas taxas como essencialmente invariáveis sob condições terrestres normais, pesquisas recentes têm sugerido possíveis influências de fatores como campos eletromagnéticos intensos, pressões extremas, e ambientes de plasma de alta energia. Estudos sistemáticos sobre estas possíveis influências, utilizando tecnologias modernas de aceleradores de partículas e instalações de plasma, poderiam refinar nossa compreensão dos limites da constância do decaimento radioativo e potencialmente identificar assinaturas geoquímicas de episódios passados de decaimento acelerado.
Outra área promissora para pesquisas futuras é o desenvolvimento de novos métodos de datação independentes que possam complementar ou verificar técnicas radiométricas existentes. Abordagens baseadas em fenômenos físicos fundamentalmente diferentes, como termoluminescência, ressonância paramagnética eletrônica, ou racemização de aminoácidos, oferecem verificações independentes de cronologias estabelecidas. A discordância sistemática entre diferentes métodos poderia indicar problemas com premissas fundamentais, enquanto concordância fortaleceria a confiança na robustez da cronologia estabelecida. Adicionalmente, o refinamento de técnicas de datação relativa, como dendrocronologia estendida, varves lacustres, e núcleos de gelo, poderia fornecer registros contínuos de alta resolução que serviriam como pontos de ancoragem cronológica independentes de pressupostos sobre constância de decaimento radioativo.
Perspectivas Futuras em Estudos de Impacto
O campo de estudos de impactos de asteroides continua a evoluir rapidamente, impulsionado por avanços tecnológicos e interesse científico crescente nas interações entre a Terra e seu ambiente cósmico. Várias direções de pesquisa promissoras emergiram recentemente, oferecendo potencial para aprofundar nossa compreensão dos efeitos de impactos na história terrestre.
Uma área particularmente dinâmica é o mapeamento sistemático e caracterização detalhada de crateras de impacto em todo o planeta. Tecnologias avançadas de sensoriamento remoto, incluindo radar de penetração no solo e imagens hiperespectrais, estão revelando estruturas de impacto anteriormente não reconhecidas, muitas delas obscurecidas por erosão, vegetação ou subsequente atividade geológica. Projetos como o Earth Impact Database estão compilando informações abrangentes sobre crateras confirmadas, enquanto estudos geofísicos de alta resolução estão revelando detalhes da estrutura subsuperficial de crateras conhecidas, permitindo reconstruções mais precisas da dinâmica de impacto e estimativas aprimoradas de idades e energias envolvidas.
Outra fronteira promissora envolve a investigação detalhada dos efeitos físicos e químicos do plasma gerado durante impactos. Experimentos em laboratório utilizando lasers de alta potência e canhões de gás para simular condições de impacto estão fornecendo insights sobre os processos de vaporização, ionização e subsequentes reações químicas que ocorrem durante estes eventos extremos. Modelagem computacional avançada complementa estes experimentos, permitindo simulações detalhadas da evolução de plasmas de impacto e seus efeitos em materiais circundantes. Estas pesquisas têm aplicações não apenas para compreender a história de impactos terrestres, mas também para mitigação de ameaças de asteroides, exploração planetária e desenvolvimento de novos materiais inspirados em processos de impacto de alta energia.
Conclusões
Esta investigação examinou as complexas interações entre impactos de asteroides e seus efeitos na geologia sedimentar, geocronologia radiométrica e DNA dos seres vivos. A análise apresentada sugere que a física extrema associada a grandes impactos – incluindo atrito intenso, efeitos piezoelétricos, temperaturas extremas, ondas sonoras e formação de plasmas de alta energia – pode ter consequências muito mais profundas e de longo alcance do que tradicionalmente reconhecido no paradigma científico atual.
A hipótese central proposta – que impactos significativos de asteroides podem ter ocorrido em um passado relativamente recente, afetando fundamentalmente os sistemas utilizados para datação radiométrica e causando mutações genéticas aceleradas – oferece um quadro interpretativo que potencialmente reconcilia diversas observações anômalas. Entre estas incluem-se a presença de tecidos moles em fósseis supostamente antigos, o paradoxo da estase morfológica no registro fóssil, a disparidade no acúmulo de mutações entre amostras antigas e contemporâneas, e a distribuição biogeográfica peculiar de espécies endêmicas.
Esta pesquisa abre um novo campo de investigação na interseção entre física nuclear, geologia e biologia evolutiva. A interdependência proposta entre temperatura, ondas sonoras, efeito piezoelétrico e formação de plasma, culminando na superação da barreira de Coulomb e aceleração do decaimento radioativo, representa uma hipótese testável que pode ser investigada através de abordagens experimentais e observacionais modernas. Estudos futuros focados nos ângulos de impacto, assinaturas químicas de famílias de asteroides, e possíveis órbitas de fragmentos relacionados oferecerão oportunidades para validar ou refutar aspectos específicos desta hipótese, contribuindo para uma compreensão mais completa da história terrestre e dos processos que moldaram sua superfície e biota.
Recomendações para Investigações Futuras
Para avançar o entendimento dos efeitos de grandes impactos de asteroides na geologia, geocronologia e evolução biológica, recomendamos várias linhas de investigação que poderiam fornecer evidências críticas para testar as hipóteses apresentadas neste estudo.
Primeiramente, sugerimos a condução de experimentos laboratoriais de física nuclear visando quantificar os efeitos de plasmas de alta energia nas taxas de decaimento radioativo. Utilizando aceleradores de partículas e instalações de plasma existentes, seria possível submeter amostras contendo isótopos radioativos a condições que simulam parcialmente aquelas geradas durante impactos. A medição precisa de alterações nas taxas de decaimento sob estas condições forneceria dados cruciais sobre a plausibilidade física da aceleração de decaimento proposta. Paralelamente, modelagem computacional avançada poderia simular as condições extremas de temperatura, pressão e ionização presentes em plasmas de impacto, permitindo previsões teóricas que complementariam os estudos experimentais.
Uma segunda abordagem essencial seria a realização de um levantamento sistemático e datação comparativa de materiais associados a crateras de impacto conhecidas. Este estudo aplicaria múltiplos métodos de datação independentes (radiométricos, termoluminescência, estratigráficos) às mesmas amostras, buscando identificar padrões consistentes de discrepância que poderiam indicar episódios de decaimento acelerado. Adicionalmente, análises detalhadas da composição isotópica de rochas dentro e ao redor de crateras poderiam revelar assinaturas de reações nucleares induzidas por impacto, como concentrações anômalas de isótopos específicos que seriam difíceis de explicar por processos geológicos convencionais. A integração destes estudos com análises de DNA antigo e taxas de mutação em populações humanas e animais poderia fornecer um quadro cronológico mais completo e potencialmente revelar correlações entre eventos de impacto e saltos no acúmulo de mutações genéticas.
Referências Bibliográficas – Parte I
- Artemieva, N., & Morgan, J. (2009). Modeling the formation of the K-Pg boundary layer. Icarus, 201(2), 768-780.
- Artemieva, N., & Shuvalov, V. (2016). From Tunguska to Chelyabinsk via Jupiter. Annual Review of Earth and Planetary Sciences, 44, 37-56.
- Armitage, M. H. (2013). Soft sheets of fibrillar bone from a fossil of the supraorbital horn of the dinosaur Triceratops horridus. Acta Histochemica, 115(6), 603-608. https://doi.org/10.1016/j.acthis.2013.01.001
- Bardeen, C. G., Garcia, R. R., Toon, O. B., & Conley, A. J. (2017). On transient climate change at the Cretaceous−Paleogene boundary due to atmospheric soot injections. Proceedings of the National Academy of Sciences, 114(36), E7415-E7424.
- Baumgardner, J. (2005). Carbon-14 Evidence for a Recent Global Flood and a Young Earth. Technical Papers of Discovery Center For Science & Earth History. www.icr.org
- Benton, M. J. (2009). The Red Queen and the Court Jester: Species diversity and the role of biotic and abiotic factors through time. Science, 323(5915), 728-732.
- Bertsch, G. F., et al. (2014). Nuclear Reactions in Astrophysics. Physical Review C.
- Berthault, G., Lalomov, A. V., & Tugarova, M. A. (2011). Reconstruction of paleolithodynamic formation conditions of Cambrian-Ordovician sandstones in the Northwestern Russian platform. Lithology and Mineral Resources, 46(1), 60-70. https://doi.org/10.1134/S0024490211010020
- Boslough, M. B., & Crawford, D. A. (2008). Low-altitude airbursts and the impact threat. International Journal of Impact Engineering, 35(12), 1441-1448.
- Bottke, W. F., et al. (2002). The origin and evolution of stony meteorites. In Asteroids III (pp. 395-408). University of Arizona Press.
- Bottke, W. F., et al. (2006). The Origin of Asteroids: A New Perspective. Nature, 439(7074), 147-151. https://doi.org/10.1038/nature04419
- Chang, J. S. (2004). Recent ancestral origins of human Y chromosomes. Nature Genetics, 36, 339-340.
- Chapman, C. R. (2004). The hazard of near-Earth asteroid impacts on earth. Earth and Planetary Science Letters, 222(1), 1-15. https://doi.org/10.1016/j.epsl.2004.03.004
- Cho, H. E., Horstemeyer, M. F., & Baumgardner, J. R. (2017). Unified Static and Dynamic Recrystallization Model for the Minerals of Earth’s Mantle Using Internal State Variable Model. AGU Fall Meeting Abstracts, 24.
- Cohen, J. S. (1988). Impact Events and Their Role in Geological Evolution. Annual Review of Earth and Planetary Sciences, 17, 207-221. https://doi.org/10.1146/annurev.ea.17.050489.001231
Referências Bibliográficas – Parte II
- Collins, G. S., Melosh, H. J., & Marcus, R. A. (2005). Earth Impact Effects Program: A Web-based computer program for calculating the regional environmental consequences of a meteoroid impact on Earth. Meteoritics & Planetary Science, 40(6), 817-840.
- Crawford, D. A., & Schultz, P. H. (2014). Electromagnetic properties of impact-generated plasma, vapor and debris. International Journal of Impact Engineering, 23(1), 169-180.
- French, B. M., & Koeberl, C. (2010). The convincing identification of terrestrial meteorite impact structures: What works, what doesn’t, and why. Earth-Science Reviews, 98(1-2), 123-170.
- Glass, B. P., & Simonson, B. M. (2013). Distal Impact Ejecta Layers: A Record of Large Impacts in Sedimentary Deposits. Springer.
- Glass, B. P., & Simonson, B. M. (2017). Meteorite impact ejecta: Mechanisms of formation and classification. In Geochimica et Cosmochimica Acta (Vol. 197, pp. 280-293).
- Glikson, A. Y., Allen, C., & Vickers, J. (2004). Multiple 3.47-Ga-old asteroid impact fallout units, Pilbara Craton, Western Australia. Earth and Planetary Science Letters, 221(1-4), 383-396. https://doi.org/10.1016/s0012-821x(04)00104-9
- Hassler, S. W., & Simonson, B. M. (2001). The Sedimentary Record of Extraterrestrial Impacts in Deep‐Shelf Environments: Evidence from the Early Precambrian. The Journal of Geology, 109(1), 1-19. https://doi.org/10.1086/317958
- Heck, P. R., et al. (2017). Rare meteorites common in the Ordovician period. Nature Astronomy, 1(2), 0035.
- Hu, J. E., et al. (2015). External Influences on Radioactive Decay. Nuclear Instruments and Methods in Physics Research.
- Johnson, B. C., & Melosh, H. J. (2012). Impact spherules as a record of an ancient heavy bombardment of Earth. Nature, 485(7396), 75-77.
- Johnson, B. C., & Melosh, H. J. (2022). The formation of the Vredefort dome: Insights from numerical modeling. Journal of Geophysical Research: Planets, 127(4), e2021JE007080.
- Jourdan, F., Reimold, W. U., & Deutsch, A. (2012). Dating Terrestrial Impact Structures. Elements, 8(1), 49-53.
- Koeberl, C., Reimold, W. U., & Shirey, S. B. (2018). Meteoritics and planetary science. Elements, 14(3), 152-153.
- Lieberman, M. A., & Lichtenberg, A. J. (2005). Principles of Plasma Discharges and Materials Processing. Wiley.
- McCoy, B. J., et al. (2013). Electron Acceleration in Plasma Waves. Physics of Plasmas.
- Melosh, H. J. (1989). Impact cratering: A geologic process. Oxford University Press.
Referências Bibliográficas – Parte III
- Melosh, H. J., & Collins, G. S. (2019). Planetary science: Meteor Crater formed by low-velocity impact. Nature, 565(7741), 596-597.
- Ormö, J., et al. (2014). First known Terrestrial Impact of a Binary Asteroid from a Main Belt Breakup Event. Scientific Reports, 4(1). https://doi.org/10.1038/srep06724
- Osinski, G. R., & Pierazzo, E. (2013). Impact cratering: Processes and products. John Wiley & Sons.
- Pierazzo, E., & Artemieva, N. (2012). Local and global environmental effects of impacts on Earth. Elements, 8(1), 55-60.
- Qin, L., & Humayun, M. (2020). Meteorites and the early Solar System. The Planetary Science Journal, 1(2), 48.
- Reimold, W. U., & Gibson, R. L. (1996). Geology and evolution of the Vredefort impact structure, South Africa. Journal of African Earth Sciences, 23(2), 125-162.
- Reimold, W. U., & Koeberl, C. (2014). Impact structures in Africa: A review. Journal of African Earth Sciences, 93, 57-175.
- Richards, M. A. (2015). Triggering of the largest Deccan eruptions by the Chicxulub impact. Geological Society of America Bulletin, 127(11-12), 1507-1520.
- Robertson, D. S., Lewis, W. M., Sheehan, P. M., & Toon, O. B. (2013). K-Pg extinction: Reevaluation of the heat-fire hypothesis. Journal of Geophysical Research: Biogeosciences, 118(1), 329-336.
- Schmitz, B., & Bowring, S. A. (2001). The Role of Extraterrestrial Impacts in the Evolution of Earth. Geology, 29(11), 1003-1006. https://doi.org/10.1130/0091-7613(2001)029<1003:TROEII>2.0.CO;2
- Simpson, G. G. (1944). Tempo and Mode in Evolution. Columbia University Press.
- Simonson, B. M., & Glass, B. P. (2004). Spherule layers—records of ancient impacts. Annual Review of Earth and Planetary Sciences, 32, 329-361.
- Sodré, G. B. (2009). Formação sedimentar entre Cambriano e Pleistoceno: indicadores de catastrofismo. Revista Brasileira de Geologia, 24(3), 78-96.
- Sodré, G. B. (2017). Ausência de desbaste em rochas impactadas por cachoeiras: Implicações para a geocronologia. Anais da Academia Brasileira de Ciências Geológicas, 12(2), 45-67.
- Sodré, G. B. (2024). Pedras pontiagudas como evidência de uma Terra jovem: Análise quantitativa de morfologias líticas. Journal of Alternative Geological Perspectives, 15(1), 112-134.
- Svetsov, V. V., & Shuvalov, V. V. (2016). Thermal radiation and luminous efficiency of superbolides. Earth and Planetary Science Letters, 435, 145-154.
- Tagle, R., & Hecht, L. (2006). Geochemical identification of projectiles in impact rocks. Meteoritics & Planetary Science, 41(11), 1721-1735.
- Tanaka, K. L., et al. (2019). Asteroid Impacts and Their Effects on Earth’s Geology. Geology.
- Toon, O. B., Bardeen, C., & Garcia, R. (2016). Designing global climate and atmospheric chemistry simulations for 1 and 10 km diameter asteroid impacts using the properties of ejecta from the K-Pg impact. Atmospheric Chemistry and Physics, 16(20), 13185-13212.
- Wang, Z. L., et al. (2016). Piezoelectric effect in materials under high-strain conditions. Journal of Applied Physics, 119(15), 154104.
- Wiegert, P. A., & Innanen, K. A. (2002). Asteroid Dynamics and Impacts. Celestial Mechanics and Dynamical Astronomy, 83(1-2), 121-133. https://doi.org/10.1023/A:1014559907551
- Wünnemann, K., Collins, G. S., & Osinski, G. R. (2008). Numerical modelling of impact melt production in porous rocks. Earth and Planetary Science Letters, 269(3-4), 530-539.
- Wünnemann, K., et al. (2016). Impact cratering in sandstone: The MEMIN pilot study on the effect of pore water. Meteoritics & Planetary Science, 51(10), 1762-1794.
- Zhang, Y., et al. (2008). Electron acceleration mechanisms in plasma. Physics of Plasmas, 15(5), 056701.
Efeitos de Grandes Impactos de Asteroides na Geologia Sedimentar, Geocronologia Radiométrica e DNA dos Seres Vivos
Este documento examina os efeitos catastróficos de impactos de asteroides na Terra, analisando suas consequências para a geologia sedimentar, a confiabilidade da geocronologia radiométrica e as mutações no DNA dos seres vivos. Apresentamos evidências científicas sobre como estes eventos extremos podem ter acelerado processos de decaimento radioativo, gerado formações geológicas extensas e causado alterações genéticas significativas nas espécies sobreviventes.